



Ⅰ. 서 론
金銀花散은 《衛生寶鑑》에 기록된 방제로 金銀花와 甘草로 구성되며 癰疽, 瘡瘍, 梅毒 등을 치료한다1). 金銀花(Lonicera Japonica)는 忍冬科에 속하는 인동덩굴의 花蕾를 말하며 淸熱解毒, 凉散風熱의 효능으로 溫病發熱, 丹毒, 癰腫疔瘡 등을 치료한다2,3). 甘草(Glycyrrhizae Radix)는 콩과의 뿌리와 뿌리줄기로 解毒, 和中緩急, 調和諸藥 등의 효능이 있어 癰疽瘡瘍, 消化性潰瘍 등을 치료한다4). 《東醫寶鑑》의 雜病篇 癰疽에서 加味芷貝散, 瓜蔞散, 橘葉散을 소개하며 金銀花, 甘草 배합을 癰疽 질환에 활용한 바 있다. 또한 仙方活命飮, 千金內消散 역시 金銀花, 甘草의 배합으로 모든 癰疽의 毒腫을 치료하며, 특히 瘡이 사지에 있을 때는 金銀花를 君藥으로 사용한다한 바 있다5). 이와 같이 金銀花와 甘草를 배오하여 염증성 질환에 다용한 점에서 착안하여 金銀花, 甘草로 구성된 金銀花散으로 연구를 진행하였다.
국내에서 金銀花 및 甘草 추출물에 대한 연구는 활발히 진행되고 있다. 주 등3)이 발표한 논문에서 金銀花가 항균, 항염증 효과가 있다고 밝혀 류마티스 관절염, 세균 감염, 항암 치료 등에 활용되고 있다고 하였으며, 박 등6)은 甘草가 세포 사멸, 세포 주기 억제 및 전이 억제 효능이 있어 항암 및 항염 치료에 사용할 수 있다고 하였다. 또한 차 등7)의 연구에서는 Dextran Sulfate Sodium으로 유도된 생쥐의 궤양성 대장염에서 金銀花 추출물의 항산화, 항염증 효과를 입증하였다. 이 등8)은 金銀花의 항염증, 항산화 작용을 보고하였으며, 윤 등9)은 설치류의 Raw 264.7 cell을 대상으로 한 실험에서 金銀花의 항염증 작용을 보고하였다. 이 등10)은 金銀花와 金銀花 전초의 혼합물이 염증과 과량의 NO, cytokine 생성과 관련된 면역 및 염증 치료에 활용될 수 있음을 보고하였다. 또한 甘草의 추출물에 대한 연구로 김 등11)의 그람 양성균에 대한 항균 효과, 김 등12)의 궤양성 대장염에서의 항염증 효과, 윤 등13)의 염증성 cytokine 억제 효과, 이 등14)의 세포 자멸사 유도 효과 등이 보고된 바 있다. 또한 이 등15)은 金銀花-甘草 복합 추출물을 이용하여 in vivo에서 궤양성 대장염에 대한 항염증 효과를 입증한 바 있다. 하지만 아직까지 金銀花散의 항염증 기전에 대한 in vitro 연구는 이루어지지 않았다.
이에 저자는 金銀花散 열수 추출물을 Raw 264.7 cell에 전처치 후 LPS로 처리하여 NO의 생성 및 iNOS의 발현, 염증성 cytokine의 생성과 NF-κB 신호 전달 체계에 어떠한 영향을 미치는지 살펴보았다.
Ⅱ. 재료 및 방법
金銀花散(Geumeunhwa-san)의 구성 약재인 金銀花, 甘草는 대원약업사(대구, 대한민국)에서 구입하여 불순물을 제거하고 grinder를 이용하여 분쇄하였다. 金銀花散 50g에 1.5L의 물을 첨가하고 대웅 약탕기로 100℃에서 3시간 동안 추출하였다(Table 1). 1차 여과를 거즈로 한 후, 여과액을 3분 동안 3,000×g에서 원심 분리 하였다. 원심 분리를 한 후 상층액만 취하여 No.2 filter paper(Clifton, NJ, USA)로 여과하였다. 이 여과액을 진공 회전 농축기(EYELA, Tokyo, Japan)로 농축하였다. 농축된 金銀花散액은 초저온 냉동기(Operon, Korea)에서 24시간 이상 동결하였으며, 최종적으로 동결 건조기를 이용하여 동결 건조물 17.56g을 얻었다. 제조한 金銀花散 열수 추출물(GEHS, Geumeunhwa-san water extract)은 사용할 때까지 4℃에서 보관하였다. 金銀花散 열수 추출물의 최종 수율은 35.12%였다. 건조된 열수 추출물은 세포 배양액인 DMEM(Dulbecco's modified eagle's medium)에 1㎎/㎖의 농도가 되도록 정량하여 용해 후 실험에 사용하였다.
| Herbal Name | Scientific Name | Amount(g) |
|---|---|---|
| 金銀花 | Lonicera Japonica | 40 |
| 甘草(炒) | Glycyrrhizae Radix | 10 |
| Total | 50 |
세포 배양을 위한 시약으로 10% FBS(fetal bovine serum) 및 DMEM, penicillin, 100㎍/㎖ streptomycin은 Gibco/BRL(Eggenstein, Germany)에서 구입하였다. MTT(3-(4,5-dimethylthiazol-2-yl)-2, 5-diphenyl-tetrazolium bromide)와 LPS(Escherichia coli 055:B5) 및 Griess reagent는 Sigma(St. Louis, MO, USA)에서 구입하였다. Dimethylsulfoxide(DMSO)은 Junsei Chemical Co., Ltd.(Tokyo, Japan)에서, Griess reagent system은 Promega(Madison, WI, USA)에서 구입하였다. Anti-iNOS의 항체는 BD biosciences(San Jose, CA, USA)에서 구입하였고, anti-NF-κB 및 anti-β-actin의 항체는 Santa Cruz Biotechnology(Santa Cruz, CA, USA)에서 구입하였으며, anti-phospho-I-κBα(p-I-κBα) 및 anti-I-κBα(I-κBα), anti-Lamin A/C의 항체는 Cell Signaling Technology(Danvers, MA, USA)에서 구입하였으며, 전기 영동 후 단백질을 전이하기 위한 nitrocellulose(NC) membrane은 Schleicher & Schuell(Dassel, Germany)에서 구입하였다.
Murine macrophage cell line인 Raw 264.7 cell은 한국세포주은행(Seoul, Korea)에서 분양받아 사용하였으며, 5% CO2, 37℃ 조건에서 DMEM 배지에 10% FBS, 100㎛/㎖ streptomycin, 100U/㎖ penicillin을 함유하여 80-90% confluence에 도달한 경우와 계대배양 횟수가 20회를 넘기지 않은 세포만을 실험에 사용하였다. FBS가 함유되지 않은 DMEM에서 Raw 264.7 cell을 추가적으로 배양한 후, 10-300㎍/㎖ 농도의 GEHS를 함유한 DMEM으로 교환하였다. GEHS를 처치한 후 각각의 실험에서 1㎍/㎖의 LPS를 사용하였다.
6 well plate에서 Raw 264.7 cell에 10-300㎍/㎖ GEHS로 처치 후 추가적으로 1㎍/㎖의 LPS를 처치하여 18시간 후 세포 배양액을 회수하였다. Nitric oxide(NO)의 생성량 측정에서 세포 배양액 내 불순물을 제거하기 위해 3,000×g에서 회수한 세포 배양액을 3분간 원심 분리 하여, 세포 배양 상등액 50㎕와 sulfanilamide, phosphoric acid 및 α-naphthylamide를 함유한 Griess reagent 50㎕를 96 well plate에 혼합하였다. 이를 차광 후 상온에서 10분간 반응시킨 후 이를 microplate reader(Model; Infinite M200 pro, Tecan, Männedorf, Switzerland)로 540㎚에서 흡광도를 측정하였다. NO의 양은 control 대비 각 군의 배수(fold)로 나타내었다.
GEHS + LPS에 대한 세포 생존율을 분석하기 위해 10-300㎍/㎖ 농도의 GEHS로 Raw 264.7 cell에 처치하고, 1시간 후에 LPS를 1㎍/㎖의 농도로 처치한 후 18시간 배양하였다. 배양 후 PBS(Phosphate buffered saline)에 MTT를 0.1㎎/㎖의 농도로 녹여서 각 well당 50㎕씩 넣고 4시간 추가배양 하였다. 배양 후 MTT 용액을 제거한 후 생성된 formazan crystal에 200㎕의 DMSO를 첨가하여 완전히 용해시키고, microplate reader(Model; Infinite M200 pro, Tecan, Männedorf, Switzerland)로 570㎚에서 흡광도를 측정하였다. 다음의 수식에 의해 상대적인 세포 생존율을 계산하여 백분율로 표시하였다.
염증 관련 cytokine을 측정하기 위해서 6 well plate에 Raw 264.7 cell을 well당 5×105 cell로 분주하여 GEHS를 100, 300㎍/㎖로 처치한 후 1시간 뒤에 다시 LPS를 처치하였다. LPS를 18시간 동안 처리한 후 세포 배양액 내 불순물을 제거하기 위해 3,000×g에서 5분간 원심 분리하여 상등액을 회수하였다. Tumor necrosis factor-alpha(TNF-α)와 interleukin (IL)-1β 및 IL-6를 각각 ELISA kit(Pierce endogen, Rockford, IL, USA)로 측정하여 각 cytokine의 함량을 측정하였다.
처치가 종료된 Raw 264.7 cell은 핵 분획 추출물(nuclear fraction)과 전세포 추출액(whole cell lysate)으로 분리하였다. Ice-cold PBS로 처치가 종료된 전세포 추출액의 세포를 세척한 후 scrapper로 cell을 수집한 뒤 microtube로 옮겼다. 여기에 radioimmunoprecipitation(RIPA) buffer(25 mM Tris-HCl pH 7.6, 150mM NaCl, 1% NP-40, 1% sodium deoxycholate, 0.1% SDS)와 Halt protease and phosphatase inhibitor cocktail(Thermo Scientific, Rockford, IL, USA)을 혼합한 lysis buffer를 첨가하여, 4℃에서 30분 동안 반응시키고 15,000×g에서 10분 동안 원심 분리하여 상등액을 취하였다. 본 상등액이 전세포 추출액으로 BCA protein assay kit를 이용하여 전세포 추출액의 단백질 함량을 정량하였다.
핵 분획은 ice-cold PBS로 세척하여 scrapper로 cell을 수집한 뒤 microtube에 옮겨 10mM Hepes(pH 7.9), 10mM KCl, 0.1mM EDTA, 0.5% Noridet P-40, 1mM dithiothreitol, 0.5mM phenylmethylsulfonyl fluoride를 포함한 완충액을 100㎕ 넣어 vortex하고 10분 동안 ice에 방치한 후 4℃에서 16,000×g로 5분 동안 원심 분리하였다. 원심 분리 후 세포질 추출액이 포함된 상등액을 제거하고 핵이 포함된 침전물에 다시 20mM Hepes(pH 7.9), 400mM NaCl, 1mM EDTA, 10mM dithiothreitol, 1mM phenylmethylsulfonyl fluoride를 함유한 완충액을 40㎕ 넣은 후 현탁시켜 ice에서 1시간 반응시키고, 4℃에서 16,000×g로 10분간 원심 분리한 후의 상등액을 핵 분획으로 얻었다. 핵 분획은 BCA protein assay kit를 이용하여 단백질 함량을 정량하였으며, 실험 시까지 –70℃에 보관하였다.
전세포 추출물 및 핵 분획의 단백질 함량을 측정하여 동일 양의 단백질을 젤 전기 영동법(SDS-PAGE)으로 전기 영동 시킨 후, NC membrane으로 단백질을 전이시켰다. 단백질이 전이된 NC membrane에 iNOS, I-κBα, p-I-κBα, NF-κB, Lamin A/C 및 β-actin의 1차 antibody와 반응시킨 후, blocking, washing과정을 거치고, horseradish peroxidase가 결합된 2차 항체와 반응시켜서 ECL detection reagents(Amersham Biosciences Corp., Piscataway, NJ, USA)로 발색한 후 Image analyzing system(Ultra-Violet Products Ltd., Upland, CA, USA)을 사용하여 단백질의 발현량을 평가하였다. 각 단백질의 발현량은 control 대비 각 실험군의 fold로 나타내었다.
Ⅲ. 결 과
NO의 생성 정도를 LPS로 활성화된 Raw 264.7 cell에서 측정하여 GEHS의 항염증 효과를 평가하였다. LPS군에서는 control군(1.00 ± 0.01)에 비교하여 NO의 생성량이 5.21 ± 0.12배 증가하여 NO의 유의성 있는 증가가 있었다. GEHS 10㎍/㎖에서는 4.72 ± 0.01배로 유의성이 없었으나, GEHS 30㎍/㎖에서는 4.21 ± 0.65배, GEHS 100, 300㎍/㎖에서는 각각 2.86 ± 0.43, 1.57 ± 0.10배로 LPS군과 비교하여 NO의 생성량이 유의하게 감소하였다(Fig. 1).
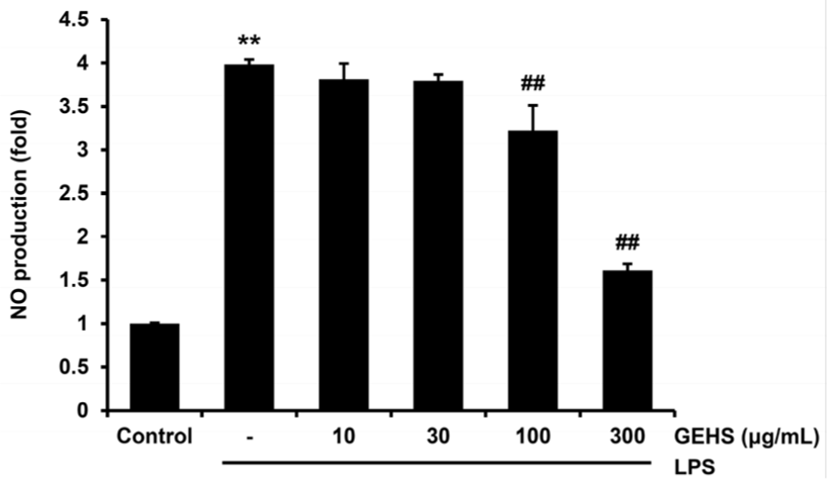
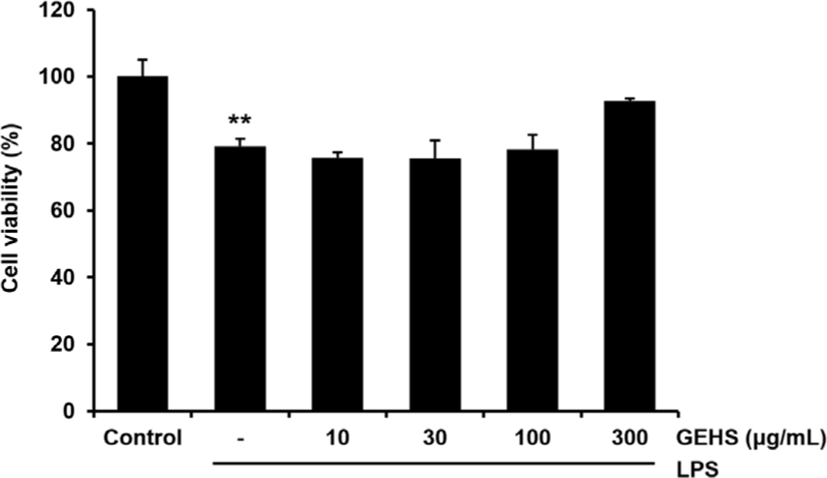
LPS에 의한 NO의 생성량은 GEHS 30, 100, 300㎍/㎖의 농도에서 유의미한 감소가 있었다. 이러한 감소의 이유가 Raw 264.7 cell에서의 GEHS와 LPS의 세포 독성 영향에 의한 것인지를 관찰하기 위해 GEHS + LPS를 처리하고 18시간 후에 MTT assay로 세포 생존율을 구하였다. LPS군에서는 control군(100.00 ± 5.10)에 비교하여 79.2 ± 2.21%의 세포 생존율을 나타내었고, LPS와 GEHS(10-300㎍/㎖)를 처리한 군에서는 세포 생존율이 각각 75.81 ± 1.62, 75.51 ± 5.48, 78.40 ± 4.21, 92.69 ± 0.84(%)로 LPS군에 비교하여 유의한 세포 생존율의 저하는 나타내지 않았다(Fig. 2). GEHS 30, 100, 300㎍/㎖의 농도에서의 NO생성 억제 결과와 GEHS(10, 30, 100, 300㎍/㎖)에서의 세포 생존율의 결과에 근거하여 이후의 GEHS의 항염증 효과 기전 연구는 GEHS 100, 300㎍/㎖의 농도에서 실험을 진행하였다.
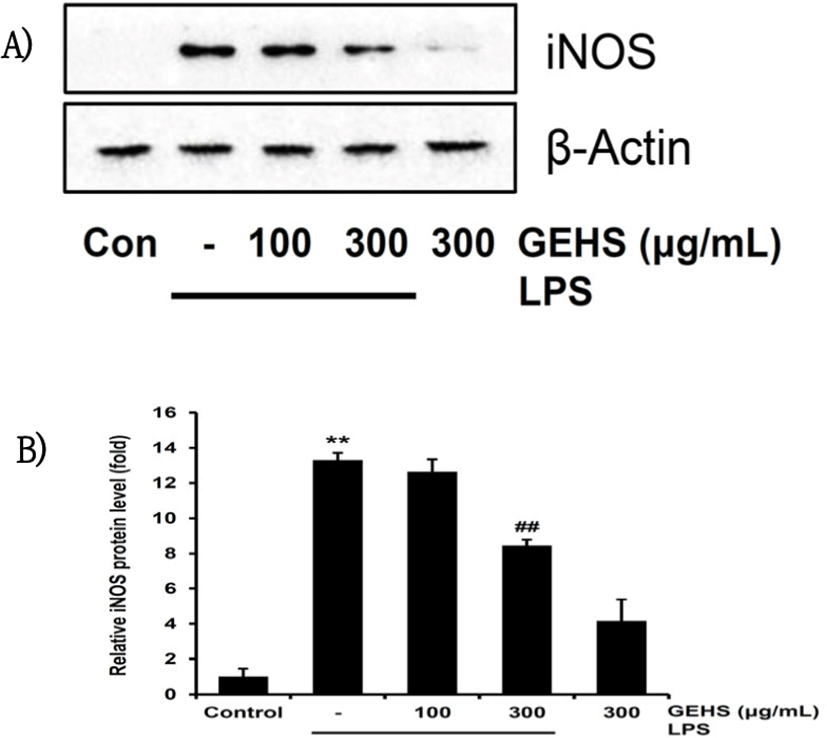
NO는 iNOS 단백질의 합성을 담당하는 핵심 효소이다. GEHS 100, 300㎍/㎖ 농도가 NO의 생성을 유의 하게 억제한 것이 GEHS의 iNOS 단백질의 조절과 관련이 있는지 확인하기 위하여, GEHS 100, 300㎍/㎖의 농도로 1시간 동안 처치한 후 Raw 264.7 cell에 1㎍/㎖의 LPS를 처치하여 western blot을 통해 iNOS 단백질 수준을 측정하였다. LPS 단독 처치 결과 무처치 대조군과 비교하여 iNOS의 발현을 control(1.00 ± 0.45) 대비 13.31 ± 0.39배로 통계적으로 유의하게 증가시켰다. GEHS 100㎍/㎖의 전처치는 iNOS 단백질의 발현을 12.63 ± 0.70배로 통계적으로 유의성 있게 감소시키지 못하였으나, GEHS 300㎍/㎖의 전처치는 8.46 ± 0.31배로 iNOS 단백질의 발현을 유의하게 감소시켰다(Fig. 3). 이를 통해 GEHS가 NO를 억제함에는 GEHS가 iNOS 발현의 억제와 연관성이 있음을 보여준다.
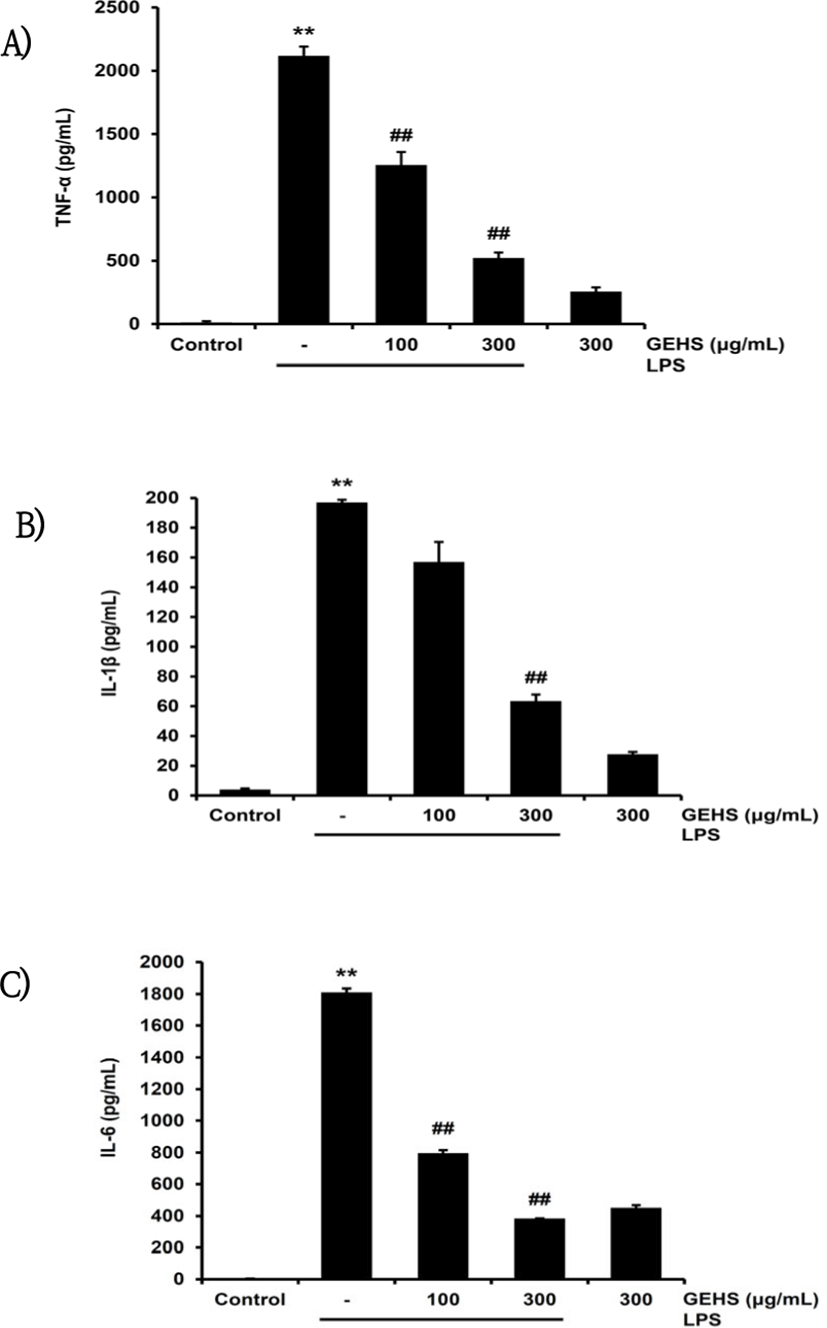
TNF-α, IL-1β, IL-6는 전 염증성 cytokine으로, LPS로 활성화된 Raw 264.7 cell에서 분비된다. 이러한 cytokine의 억제에 GEHS가 미치는 영향을 확인하기 위하여 100, 300㎍/㎖ 농도의 GEHS를 1시간 동안 전처치한 후 LPS를 처치하여 TNF-α, IL-1β, IL-6의 생성량을 측정하였다. TNF-α는 LPS에 의해 2118.44 ± 72.00pg/㎖로 무처치 대조군(12.18 ± 10.22pg/㎖)에 비해 증가함이 유의하였으나, 100, 300㎍/㎖ 농도의 GEHS 전처리에서의 TNF-α 생성량은 1256.03 ± 103.49pg/㎖, 522.22 ± 43.03pg/㎖로 LPS 단독 처치군에 비하여 유의하게 감소시켰다(Fig. 4A). IL-1β는 LPS를 단독으로 처치 시 196.9 ± 1.86pg/㎖로, 무처치 대조군(4.04 ± 0.65pg/㎖)에 비해 증가함이 유의하였다. 반면 GEHS 100, 300㎍/㎖의 전처치로 IL-1β 생성량은 각각 157.1 ± 13.36pg/㎖, 63.46 ± 4.41pg/㎖로, GEHS 300㎍/㎖에서 LPS 단독 처치군에 비해 감소함이 유의하였다(Fig. 4B). IL-6는 LPS에 의해 1810.79 ± 24.31pg/㎖로 무처치 대조군(4.75 ± 0.22pg/㎖)에 비하여 유의하게 증가하였다. GEHS 100, 300㎍/㎖을 전처치 후 IL-6 생성량은 각각 796.33 ± 18.85pg/㎖, 383.93 ± 1.52pg/㎖로, LPS 단독 처치와 비교하여 감소함이 통계적으로 유의하였다(Fig. 4C). 이를 통해 GEHS 100, 300㎍/㎖의 농도에서 LPS로 유발되는 cytokine 생성을 억제할 수 있음을 확인하였다.
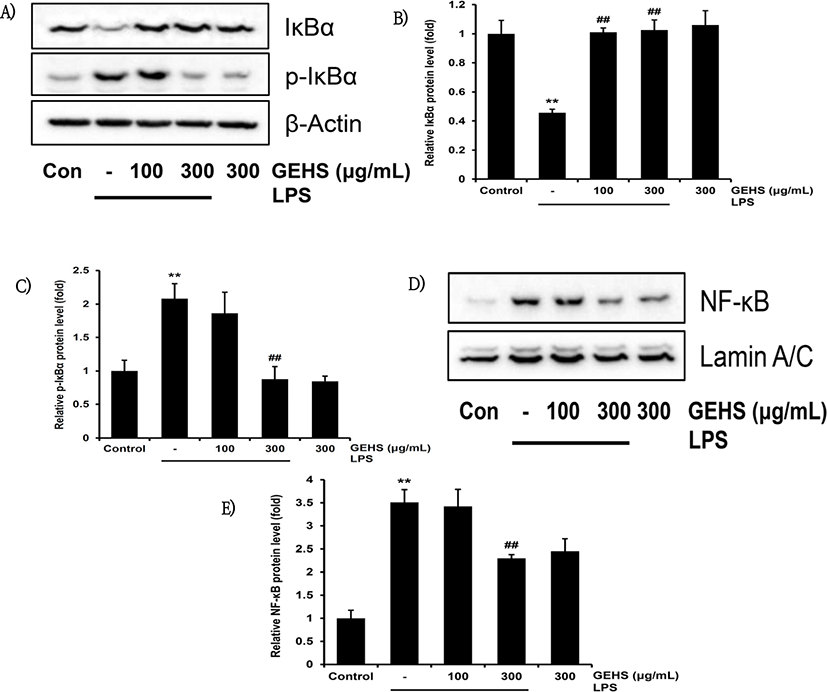
IκBα는 세포질 내 NF-κB 억제 단백질로, NF-κB의 신호 회로 활성화에 영향을 미친다16). IκBα가 인산화 되면 IκBα로부터 NF-κB가 분리되어 세포핵 안으로 이동하면서 NF-κB 신호 회로가 활성화된다16). NF-κB 신호 회로의 조절에 의해 GEHS의 염증 억제가 이루어지는 것인지 확인하기 위하여 100, 300㎍/㎖의 농도의 GEHS를 1시간 동안 전처치하고 1㎍/㎖의 LPS를 30분 동안 처치한 후 western blot으로 IκBα의 수준을 세포질에서 측정하였다. 또한 세포핵 안에서의 NF-κB 단백질 발현 및 IκBα, IκBα의 인산화 된 형태인 p-IκBα의 수준을 측정하였다. GEHS를 100, 300㎍/㎖로 전처치하였을 때 IκBα 단백질 생성량은 LPS 단독 처치(0.45 ± 0.02배)와 비교하여 각각 1.01 ± 0.03, 1.02 ± 0.06배로 통계적으로 유의하게 증가하였다(Fig. 5A, B). LPS에 의해 p-IκBα의 발현이 유의하게 증가되었으며, GEHS 100㎍/㎖으로 전처치했을 때 LPS 단독 처치군(2.08 ± 0.22배)과 비교하여 p-IκBα의 발현 수준이 1.86 ± 0.31배로 유의한 결과를 나타내지 않았다. 반면 300㎍/㎖ 농도의 GEHS을 전처치하였을 때, LPS 단독 처치군(2.08 ± 0.22배)에 비해 p-IκBα의 발현 수준이 0.87 ± 0.18배로 유의하게 감소하였다(Fig 5A, C). 또한 LPS 단독 처치 시 무처치 대조군에 비하여 NF-κB 단백질은 3.50 ± 0.27배로 유의하게 증가하였으나, LPS 단독 처치군에 비해 GEHS 100㎍/㎖의 전처치 후 LPS로 활성화하였을 때 3.42 ± 0.37배로 유의한 결과를 나타내지 않았다. 반면 GEHS를 300㎍/㎖으로 전처치 후 LPS로 활성화하였을 때 LPS 단독 처치군에 비해 2.29 ± 0.07배 유의하게 감소하였다(Fig. 5D, E).
Ⅳ. 고 찰
金銀花散은 《衛生寶鑑》1)에서 癰疽惡瘡을 主治하며 一切內外癰疽을 치료한다고 소개되어 있으며, 특히 腸癰 치료에 효과가 높다고 하여15) 염증성 질환에 대한 활용도가 높은 처방이다. ≪醫方集解≫에서 金銀花酒로 기재되어 있으며, 金銀花酒는 婦人乳癰, 一切癰疽, 背癰, 癰疽惡瘡에 사용하며 특히 腸癰에 효과가 뛰어나다고 소개되어 있고15), ≪東醫治療經驗集成≫에서는 金銀花-甘草 복합 추출물이 대장의 궤양과 미란을 호전시켜 만성 궤양성 대장염 치료제로 사용할 수 있음을 보여주었다15). 또한 ≪東醫寶鑑≫5)에서는 癰疽로 인한 열독이 성하면 심한 갈증 즉, 煩渴이 일어나는데 이를 金銀花散이 치료한다고 하였다.
국내에서도 구성 약물인 金銀花, 甘草에 관한 항염증 효과는 주 등3), 이 등8), 이 등10), 차 등7), 윤 등9), 김 등11), 이 등12), 윤 등13), 이 등14), 이 등15) 에 의해 연구된 바 있으나, 복합제제인 金銀花散에 대한 항염증 효과의 기전 연구는 이루어지지 않았다. 이 등15)의 연구에서는 金銀花-甘草 복합 추출물이 궤양성 대장염의 염증을 억제시킬 수 있는 가능성에 대해 제시하였고, 실험을 통해 金銀花-甘草 복합 추출물은 항염증 효과 및 창자 점막 보호 효과가 있음을 밝혔다. 하지만 이는 in vivo에서 얻은 결과로, 추가적인 실험 자료가 필요하였다. 이에 본 연구에서는 金銀花散의 항염증 효과 및 그 기전을 in vitro에서 연구하였다.
본 연구에서 사용한 Raw 264.7 cell은 대부분의 조직에서 상주하는 mouse의 대식세포주이다. 이는 전염증 효소인 iNOS와 COX-2를 생산하여 T림프구를 활성화시켜 염증과 면역 반응 조절에 중요한 항원 전달 세포로서의 역할을 수행한다7,17). 또한 염증 반응을 유발하는 인자로 LPS를 사용하였다. LPS는 그람 음성 세균의 세포벽 구성 성분으로18), TLR4와 결합하여 TNF-α, IL-1, IL-6, IL-8 등을 포함한 다양한 염증성 cytokine을 생성시켜 염증 반응 연구에 빈용된다19). 염증 반응 과정에서 iNOS에 의한 과다한 NO의 생산은 산화 스트레스로 인한 조직 손상을 일으킬 수 있다. 염증 질환에 있어서 중요한 역할을 하는 NO는 염증 반응 과정에서 과량이 만들어진다. 그러므로 NO의 발생을 억제시키는 것이 염증 반응의 억제와 치료에 중요하다19).
본 연구에서는 GEHS의 항산화, 항염증 효과 여부를 확인하기 위해 먼저 NO 생성량과 세포 생존율을 연구하였다. 그 결과 GEHS과 LPS의 병용 처치는 LPS 단독 처치군에 비교하여 세포 독성을 유발하지 않았으며, GEHS 전처리 후 LPS로 염증 반응을 일으켰을 때 GEHS 30, 100, 300㎍/㎖의 농도에서 NO 생성량이 감소하였다. 또한 GEHS를 단독으로 처치한 군과 비교하여 GEHS 10–300㎍/㎖의 농도에서 LPS군에 비교하여 유의한 세포 생존율의 저하는 나타내지 않았다. 따라서 GEHS의 처리가 LPS로 인한 NO 생성을 억제하므로 세포 독성에 대한 방어 효과가 있음을 확인하였다.
다음으로 iNOS 단백질의 발현 수준을 확인하여 GEHS의 전처리 후 NO의 생성량 감소가 iNOS의 발현 감소에 인한 것임을 규명하고자 하였다. 그 결과 300㎍/㎖ 농도의 GEHS을 전처치 하였을 때 iNOS의 감소가 유의미함을 확인하여, NO의 생성량 감소가 GEHS의 iNOS 단백질 발현 억제로 인함을 확인하였다.
또한 이후의 실험은 염증 매개 인자들을 GEHS 100, 300㎍/㎖의 농도에서 확인하였다. 본 연구에서는 염증 매개 인자인 IL-1β, IL-6, TNF-α 수치를 측정하여 GEHS의 염증 반응 억제 효과를 확인하고자 하였다. 항원에 의해 활성화된 T cell, NK cell, mast cell 등에서 TNF가 분비되며, 활성화된 mononuclear phagocyte에서는 다량 분비된다. 대식세포에서 TNF의 분비를 활성화시키는 강력한 자극제가 LPS이다. TNF는 endothelial cell, neutrophil을 활성화시키고, hypothalamus에 작용하여 발열을 유도하며 세포의 apoptosis를 유도한다20).
IL-1은 TNF와 유사하게 숙주의 염증 반응에 주된 매개체로 작용하며, 활성화된 mononuclear phagocyte에서 다량 분비된다. 또한 IL-1은 endothelial cell을 활성화시키고, hypothalamus에 작용하여 발열을 유도한다. IL-1은 IL-1α와 IL-1β의 2가지가 있으며, 체내에 순환하는 IL-1의 대부분이 IL-1β이다20).
한편 IL-6는 주로 macrophage, endothelial cell, T cell에서 분비되며, B cell을 활성화하여 항체의 생성을 촉진한다20). 본 실험에서 GEHS가 세포의 신호 분자 제어를 통해 염증 반응을 억제함이 유의한지 연구한 결과, 100, 300㎍/㎖ 농도의 GEHS 전처리는 LPS로 유발되는 염증 매개 인자들의 생성을 억제할 수 있음을 밝혔다.
한편 NF-κB, AP-1 등은 iNOS, COX-2의 발현에 관련된 전사 조절 인자로서, NF-κB가 대표적이다21). NF-κB는 cell apoptosis의 억제, cell regulation, oncogenesis 등과 관련이 있으며 염증 반응에 의해 활성화되어 TNF-α나 iNOS의 발현에 관련된다22). 비활성상태의 NF-κB는 I-κB와 세포질에서 결합하여 비활성형으로 존재하다가 LPS에 의해 활성화되고, I-κB가 인산화되면서 핵으로 전위한 NF-κB는 iNOS, COX-2 등의 전사를 유도한다23). GEHS의 전처치는 세포질에서 LPS에 의해 증가된 I-κBα의 발현을 감소시키고, 핵 분획에서의 NF-κB는 GEHS의 전처치에 의해 NF-κB량이 감소하였다. 이는 I-κBα의 인산화 억제 및 NF-κB의 핵전위 억제에 GEHS가 관여하여 결과적으로 iNOS를 억제함을 의미한다.
또 PAMPs (pathogen-associated molecualr patterns)는 면역 반응을 유도하는 대식세포, 수지상세포 등 세균, 진균, 기생충, 바이러스 입자 등의 표면에 존재하는 분자다. 또한 PRRs (Pattern recognition receptors)란 이를 인식하여 면역과 관련된 유전자 발현을 촉진하는 신호 전달 경로를 활성화시키는 수용체를 일컫는다24). Toll 유사 수용체 (TLR)은 가장 먼저 발견된 PRR로, 그 중 TLR4는 그람 음성 세균, 곰팡이 등의 미생물 ligand를 인지하여 면역 반응을 일으킨다25). 본 연구에서 염증 반응 유도에 사용된 LPS는 그람 음성 세균의 세포벽 구성 성분이다. 이는 TLR4/4와 결합하여 NF-κB 회로의 활성화 및 세포 내 신호 전달 체계의 활성화로 결과적으로 전염증성 인자인 IL-1, IL-6, IL-8, TNF-α 등의 합성과 방출을 유도한다26). 또한 MyD88-의존적 신호 전달이라는 경로는 TLR과 PAMP의 결합으로 활성화된다. TLR과 해당 ligand가 결합하면 TAK1이 MyD88을 통해 활성화되고, 또 IκB 인산화(IKK) 복합체는 NF-κB와 결합되어 있다가 TAK1에 의해 인산화 된다. NF-κB는 인산화 된 IKK 복합체에서 유리되며, 염증 매개 인자의 발현은 유리된 NF-κB가 핵으로 이동함에 따라 이루어진다. 또한 MAPK 경로는 TAK1이 IKK 복합체로부터 분리된 이후 활성화되어 Fos 및 Jun의 전사가 유도된다7,25,26).
본 실험에서는 NF-κB의 전사 조절이 GEHS가 IKK 복합체의 인산화를 억제하기 때문임을 확인하고자 하였다. GEHS 300㎍/㎖를 전처치 후 LPS로 활성화 하였을 때, LPS 단독 처치군에 비교한 IκBα 단백질 생성량의 증가가 유의하였고, p-IκBα는 GEHS 300㎍/㎖ 농도에서 유의하게 감소하였으며, 세포핵 내의 NF-κB는 GEHS의 300㎍/㎖ 처리 시 유의하게 감소하였다. 결과적으로 GEHS는 IκBα의 인산화를 억제하여 NF-κB의 전사 및 염증 매개 인자의 발현을 억제함을 밝혔다.
결론적으로 본 연구는 염증성 질환에 사용하는 金銀花散(GEHS)에 대한 대식세포 수준에서 항염증, 항산화 효과 및 기전을 실험하여 GEHS가 임상적으로 활용될 수 있는 근거를 제공하였다는 점에 의의가 있다. 또한 金銀花가 한방에서 淸熱解毒, 凉散風熱 등의 효능이 있어 염증성 질환, 부스럼, 발열 등에 사용할 수 있다고 하였으며, 甘草는 解毒, 和中緩急, 調和諸藥 등의 효능으로 발열, 소화성 궤양, 식중독 등의 염증성 질환에 효과가 있다고 알려져 있는 것을 바탕으로, 金銀花-甘草의 복합 처방 역시 항염증 효과가 있음을 in vitro에서 증명하였다. 하지만 위에서 언급한 MAPK 경로 등과 같이 NF-κB 경로 외에도 염증 매개 인자의 발현에 관여하는 다른 경로가 존재하므로, GEHS의 염증 억제 기전을 규명하기 위해 추가적인 연구가 필요하며, 金銀花-甘草 배합에 따른 항염증 효과에서 金銀花散 방제의 구성 비율(金銀花:甘草 = 4:1)이 최적 비율인지는 추가적인 연구가 필요할 것으로 사료된다. 또한 본 연구는 대식세포 수준에서 이루어진 연구로, 동물 모델에서의 염증 억제 효과가 세포 수준에서와 같이 NF-κB 경로를 경유하는가에 대한 후속 연구가 필요하며, 실제 임상 치료에 적용하기 위한 다양한 염증성 질환들에 대한 金銀花散의 항염증 효과에 대한 연구가 추가적으로 필요할 것으로 생각된다.
Ⅴ. 결 론
본 연구에서는 대식세포주 Raw 264.7 cell을 LPS로 활성화시켜 염증 반응을 유도하여 GEHS의 항염증 기전을 연구하여 다음과 같은 결과를 도출하였다.
-
LPS는 NO의 생성량을 유의하게 증가시켰고, GEHS 30, 100, 300㎍/㎖의 농도에서 증가된 NO를 유의하게 억제하였다. 또한 GEHS + LPS는 LPS 단독 처리군에 비교하여 세포 독성을 나타내지 않았다.
-
LPS는 iNOS 단백질의 발현을 유의하게 증가시켰으며, GEHS는 300㎍/㎖의 농도에서 증가된 iNOS 단백질의 발현을 유의하게 감소시켰다.
-
LPS는 IL-1β, IL-6, TNF-α의 전염증성 cytokine을 유의하게 증가시켰으며, GEHS 100, 300㎍/㎖의 농도에서 증가된 IL-1β, IL-6, TNF-α의 농도를 유의하게 감소시켰다.
-
LPS는 p-IκBα 단백질의 발현을 유의하게 증가시켰고, GEHS는 300㎍/㎖의 농도에서 증가된 p-IκBα 단백질의 발현을 유의하게 감소시켰다. 또한 LPS에 의해 증가된 핵분획에서의 NF-κB 발현이 GEHS 300㎍/㎖의 농도에서 증가된 NF-κB의 발현을 유의하게 감소시켰다.
이와 같은 결과로 GEHS는 IκBα의 인산화를 억제하고, 또한 NF-κB의 핵으로의 전위를 억제하여, 항염증 효과를 나타냄을 알 수 있었다.