



Ⅰ. 서 론
최근 생활 습관 및 환경이 변화되면서 각종 유해물질이나 자외선의 노출 기회가 빈번해짐에 따라 전 세계적으로 피부암 발생빈도가 증가되었고, 피부암은 공중보건의 심각한 문제를 야기하고 있다1). 그 중 흑색종은 멜라닌 세포가 존재하는 어느 부위에나 발생할 수 있으며, 다른 피부암과 달리 성장 속도가 매우 빨라 발생 초기에 다른 장기로 전이율이 높아 악성도가 높은 종양으로 알려져 있다2). 또한 국소재발이 높은 빈도로 일어나고, 화학요법이나 방사선치료에도 저항성이 강하여 치료가 가장 어려운 악성종양 중의 하나이다3). 지금까지의 항암물질에 대한 연구는 암세포를 사멸시키는 단편적인 결과물에 치중되어 왔으나, 이에 대한 분자생물학적 수준에서의 기작 연구가 불충분한 것이 현실이다. 따라서 항암효과가 기대되는 천연물에 대한 명확한 기작 연구가 진행되어야 한다4).
서양의학에서 피부암의 치료는 외과적 절제술이 가장 일반적인 치료 방법이며 동결요법, 소파술, 전기건조법, 광역동치료, 방사선요법, 화학요법, 면역요법 등의 다양한 치료방법이 있다5). 그러나 흑색종이 다른 피부암보다 더 치명적인 이유는 항암화학요법에 대한 저항성을 가지고 있어 20% 정도의 환자에서만 치료효능을 보이기 때문이다6). 약물 저항성에 대한 기전연구는 아직 밝혀지지 않았지만 세포의 죽음 기전과 연관되어 있다고 생각되고 있으며, 또한 이것이 질병의 진행과 전이에 연관되어 있다고 알려져 있어 세포의 죽음에 관한 기전연구가 절실히 요구되고 있다7,8).
세포는 세포가 mitosis하는 시기(M phase)와 염색체 DNA를 synthesis하는 S phase, 그리고 이들의 준비를 행하는 시기(G1 and G2 phase)를 반복하여 증식하는데, 이 반복을 세포주기라 한다9). 세포의 증식과 고사는 세포주기에 의해 결정되는데, 세포주기는 regulatory cyclins subunits, cyclin dependent kinases 그리고 their inhibitors 같은 세포주기 조절인자에 의해 조절된다10). 암이라는 것은 정상적으로 세포활동이 유지되지 못하고 성장과 분열이 제어가 되지 않아서 무절제한 방식으로 세포가 증식하는 현상을 말하는데, 이러한 세포주기 제어능력을 확인하는 것이 항암활성을 검증하는 지표로 사용된다11).
와송은 돌나무과(Crassulaceae)에 속한 다년생 육질초본인 바위솔(Orostachys japonica Berger)의 전초로 여름, 가을에 채취하여 쇄건하여 사용한다. 性味는 凉酸苦하여 肝, 肺經에 歸入하며, 주로 凉血止血, 淸熱解毒, 收濕斂瘡 하는 효능이 있어 吐血, 衄血, 便血, 血痢, 熱淋, 月經不調 및 疔瘡癰腫 등의 병증에 적용한다12). 와송에는 Friedelin, epi-friedlanol, grutinone, glutinol, triterpenid, kaempferol, quercetin, flavonoidβ-sitosterol, cam-pesterol, fatty acid ester, quercetin, flavonoid 등 다양한 약리적 성분이 존재하는 것으로 밝혀졌으며13,14), 전통적으로 소염제, 해열제, 지혈제, 해독제 및 항암의 목적으로 사용되었다고 알려져 있다15). 또한 항균16), 항당뇨17) 효능과 위암15), 간암18), 대장암19) 등 여러 질환에서 유효한 작용을 나타내는 것으로 밝혀졌다. 이전 연구들에서는 와송의 생리 활성 물질이 여러 종류의 암에서 항암효과가 있다고 알려져 왔지만, 아직까지 와송이 흑색종에 미치는 세포사멸과 항암활성에 관한 연구는 없다.
이에 본 연구에서는 와송 유래 생리활성물질에 의한 인체 흑색종 A375SM 세포에 대한 항암 효과를 세포주기 조절의 관점에서 확인하고 이의 분자적 기전을 밝히고자 한다.
Ⅱ. 재료 및 방법
A375 human melanoma cell lines은 Korean cell line bank(KCLB, Seoul, Korea)로부터 구입하여 사용하였다. 세포배양과정에서는 Dulbecco’s modified Eagle’s medium(DMEM; Corning, Manassas. VA, USA), fetal bovine serum(FBS; Corning, Manassas. VA, USA), penicillin-streptomycin antibiotic solution(Hyclone, Logan, UT, USA)을 사용하였다. MTS(3-[4,5-dimethylythiazol-2-yl]-5-[3-carbpxy-methoxyphenyl]-2-[4-sulfophenyl]-2H-tetrazolium, innersalt) 용액인 CellTier 96 non-radioactive cell proliferation assay kit(Promega, Madison, WI, USA)를 이용하였고, Apoptosis 유도능과 cell cycle arrest 정도를 확인하기 위하여 MUSE® Annexin V and Dead Cell Assay Kit(Merck KGaA, Darmstadt, Germany)와 MUSE® Cell Cycle Kit(Merck KGaA, Darmstadt, Germany)를 사용하였다. 또한 4,6-diamidino-2-phenylindole(DAPI) solution(VECTASHIELD mounting medium with DAPI, Burlingame, CA, USA)으로 핵을 염색하여 형광현미경으로 확인하였다. Western blotting을 위하여 사용한 primary antibodies인 NF-κB, Bcl-2, pro-caspase-3, cleaved caspase-3, CDK1, CDK4, Cyclin B1, Cyclin D1, p-ERK, p-JNK, p-p38, ERK, JNK, p38은 cell signaling technology (Danver, MA, USA)로부터 구입하여 사용하였고, Glyceraldehyde 3-phosphate dehydrogenase (GAPDH)는 AbFrontier(Seoul, Korea)에서 구입하여 사용하였다. 그리고 Mouse anti-Rabbit IgG-HRP conjugated secondary antibodies는 Santa Cruz Biotechnology(Dallas, TX, USA)에서 구입하였다.
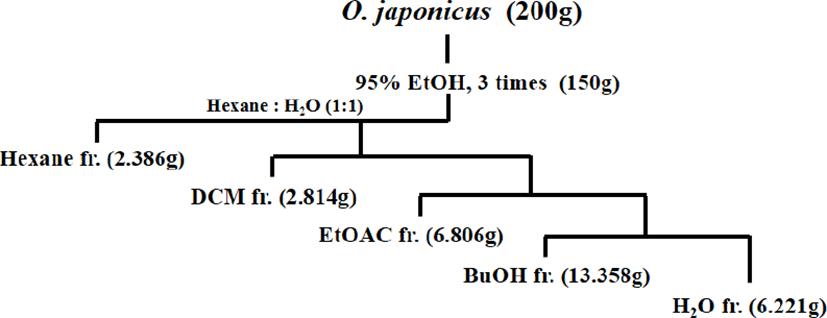
O. japonicus은 Geobugiwasong 농장(Miryang, Korea)으로부터 제공받았다. O. japonicus는 자연건조시켜, 믹서기를 이용하여 잘게 분쇄하여 powder로 만들었다. Round bottom flask안에 200g의 powder O. japonicus과 95% ethyl alcohol(EtOH) 1ℓ를 현탁한 다음 reflux condenser(SciLab®, Seoul, Korea)를 이용하여 3h, 3회 끓였다. Crude O. japonicus extract를 rotary evaporator(IKA-Werke GmbH & Co. KG, Staufen, Germany)를 사용하여 40℃에서 EtOH를 완전히 증발시켰다. 동일 플라스크에 1ℓ의 double distilled water(H2O)를 넣고 충분히 흔들어 준 후, 동량의 n-hexane(hexane), dichloromethane (DCM), ethylacetate(EtOAc), n-butanol(BuOH), H2O순으로 systematic solvent fractionation을 수행하였다(Fig. 1). 각각 회수된 solvent fraction은 rotary evaporator을 사용하여 용매를 완전히 제거하고, 유리용기에 옮겨담은 후, silica gel이 들어있는 밀폐된 용기에서 충분히 건조시키고 가루로 분쇄하여 –20℃ 냉동보관하였다. 본 연구에 사용한 용매 분획물은 이전의 연구에서 항암효과가 가장 뛰어난 것으로 밝혀진15) EtOAc 분획물로서 실험 전 세포실험전용 0.1% dimethyl sulfoxide(DMSO; applichem GmbH, Darmstadt, Germany) 용액에 녹여서 사용하였다.
본 연구에 사용한 세포주는 A375 human melanoma cell로서 10% FBS, 1% penicillin-streptomycin이 들어있는 DMEM 배지를 이용하여 37℃, 5% CO2 incubator에서 배양하였다. 세포가 90-95% confluency를 보일 때, 4-5일마다 계대배양하였고, 1-2일마다 1×PBS를 사용하여 2번 세척하고 배지를 교체해 주었다. 이때 10% trypsin(Corning, Manassas. VA, USA)을 이용하여 Dish에 부착된 세포주를 떼어내었다. 세포주는 모든 실험에서 5×105cells/㎖ 농도로 사용하였고, 농도별 EtOAc fraction(0, 20, 40, 60, 80, 100㎍/㎖)을 처리하기 전, 1% penicillin-streptomycin 만 들어 있는 serum free(SF) media를 이용하여 6h 이상 starvation 하였다.
A375 human melanoma cells에 대한 EtOAc fraction의 농도별, 시간별 세포생존율을 확인하기 위하여 CellTier 96 non-radioactive cell proliferation assay kit를 사용하였고, 실험방법은 제조사에서 지시하는 대로 수행하였다. 96 well plates에 5×105cells/㎖ 농도로 세포를 분주한 후, 18h 37℃, 5% CO2 incubator에서 배양하였다. 배양 후 SF media로 washing하고, 6h starvation 시킨 후, EtOAc fraction을 농도별(0, 20, 40, 60, 80, 100㎍/㎖)로 처리하였다. 시간별 6, 12, 24h 배양 후, 상층액을 모두 제거하고 MTS 시약 100㎕를 분주하였다. 37℃, 5% CO2 incubator에서 1h 배양 후, 470㎚에서 Multiskan sky microplate spectrophotometer (ThermoFisher Scientific, Waltham, MA, USA)를 이용하여 흡광도를 측정하였다. MTS를 이용한 cell viability assay는 3번 반복 실험하여 평균결과 및 standard deviation 수치를 산출하였다.
OJEF의 A375 human melanoma cells에 대한 cell cycle arrest 유도능을 확인하기 위하여 MUSE™ analyzer를 이용하여 G0/G1, S, G2/M phase의 %를 각각 측정하였다. A375 human melanoma cells에 대한 EtOAc fraction의 농도별 세포주기 제어능을 확인하기 위하여 MUSE® Cell Cycle Kit를 사용하였고, 실험방법은 제조사에서 지시하는데로 수행하였다. 6 well plates에 5×105 cells/㎖ 농도로 세포를 분주한 후, 18h 37℃, 5% CO2 incubator에서 배양하였다. 배양 후 SF media로 한번 washing하고, 6h starvation 시킨 후, EtOAc fraction을 농도별(0, 20, 40, 60, 80, 100㎍/㎖)로 처리하였다. 6h 배양 후, 상층액을 모두 제거하고, 1×PBS를 사용하여 2번 세척하였다. 그리고 trypsin 100㎕를 분주하여 plate에 부착된 세포를 분리하였고, 1.5㎖ e-tube에 세포를 회수하였다. 회수한 세포는 5,000rpm에서 5분간 원심분리를 수행하여 cell pellets를 수집하였고, 수집한 cell pellets은 70% cold EtOH에 현탁하여 4h이상 -20℃ 냉동고에서 fixation 시켰다. MUSE™ analyzer를 이용하여 분석하기 직전에 EtOAc 분획물의 농도별 시료를 전처리 하였다. 먼저 5000rpm에서 5분간 원심분리를 수행하여 EtOH이 남지 않도록 상층액을 제거하고, 1×cold PBS로 세척하고 1×cold PBS 100㎕에 현탁 한 후, cell cycle-MUSE solution 100㎕ 추가하고 어두운 실온에서 30분간 incubator 시켰다. 대조군 및 실험군에 대한 농도별 cell cycle arrest정도를 G1, S, G2/M phase로 구분지어서 MUSE™ analyzer(Merck KGaA, Darmstadt, Germany)을 이용하여 측정하였다. MUSE™ analyzer를 이용한 cell cycle analysis는 3번 반복 실험하여 평균결과 및 standard deviation 수치를 산출하였다.
A375 human melanoma cells에 대한 EtOAc fraction의 농도별, 시간별 단백질 발현정도를 확인하기 위하여 Western blotting을 수행하였다. 6 well plates에 5×105 cells/㎖ 농도로 세포를 분주한 후, 18h 37℃, 5% CO2 incubator에서 배양하였다. 배양 후 SF media로 한번 washing하고, 6h starvation 시킨 후, EtOAc fraction을 농도별(0, 20, 40, 60, 80, 100㎍/㎖)로 처리하였다. 3, 12, 24h 배양 후, 상층액을 모두 제거하고, cold 1×PBS를 사용하여 2번 세척하였다. Lysis buffer(Cell signaling technology, Danver, MA, USA)를 100㎕를 분주하고 scrapper을 이용하여 세포를 회수하였다. 회수한 세포는 ice에서 30분간 incubation 시킨 후 단백질 정량은 BCA시약(Pierce, Rockford, IL, USA)를 이용하여 농도를 정량한 후, 8-15% SDS-polyacrylamide gel에 전기영동(Bio-Rad, Philadelphia, PA, USA) 으로 분리하였다. 전기영동 후 단백질을 PVDF membrane(Bio-Rad, CA, USA)으로 transfer하고, membrane은 5% nonfat milk로 2h blocking 하였으며, 1:1,000으로 희석한 1차 antibody와 4℃에서 18h 이상 반응시켰다. 반응 membrane을 1×PBST buffer에 5분간 3번 세척하고, 세척 후 membrane은 1:500으로 희석한 2차 antibody와 4℃에서 2h 반응시키고 다시 1×PBST에 세 번 세척하였다. 표적 단백질은 ECL detection kit(Santa Cruz, CA, USA)를 이용하여 X-ray film에 감광시켜 확인하였고, band는 ImageJ software를 이용하여 분석하였다.
Ⅲ. 결 과
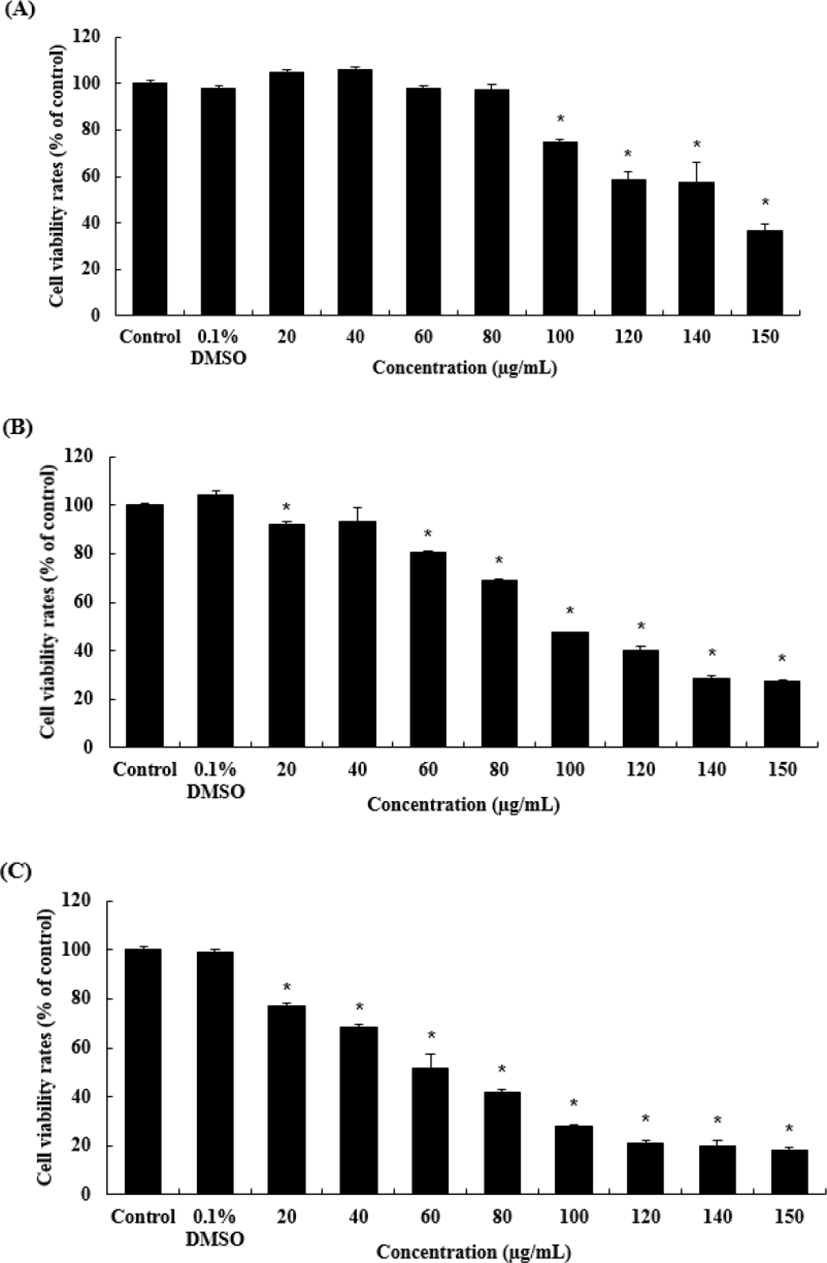
A375 세포에 대한 암세포 증식 억제효과를 MTS assay를 통하여 확인하였다. 암세포의 증식이 OJEF의 처리농도와 배양시간에 의존적으로 감소하는 것을 알 수 있었다(Fig. 2). OJEF를 농도별(0, 0.1%DMSO, 20, 40, 60, 80, 100, 120, 140, 150㎍/㎖)로 처리 하고, 12h 후 회수하여 보라색의 formazan 형성정도를 470㎚ 흡광도에서 측정하고, 대조군을 100% 생존율로 보고 각 농도별 실험군의 결과 값을 %으로 환산한 값을 그래프로 나타내었다(Fig. 2A). 그 결과, Control에 비해 각 농도별(20, 40, 60, 80, 100, 120, 140, 150㎍/㎖) 결과 값은 순서대로 104.88±1.38, 105.95±1.46, 98.05±0.90, 97.1±2.42, 74.59±1.24, 58.32±3.82, 57.39±8.58, 36.49±2.93이었고, 100㎍/㎖에서부터 유의적으로 감소하였다. OJEF를 농도별로 처리 하고, 24h 후 회수하여 MTS assay를 수행한 결과, 농도별(20, 40, 60, 80, 100, 120, 140, 150㎍/㎖)로 92.21±0.85, 93.55±5.65, 80.32±0.93, 68.78±0.80, 47.34±0.12, 39.88±1.78, 28.67±0.88, 27.47±0.37이었고, 이중 20㎍/㎖와 60㎍/㎖에서부터 유의적으로 감소하였다(Fig. 2B). OJEF를 농도별로 처리 하고, 48h 후 회수하여 MTS assay를 수행한 결과, 농도별 (20, 40, 60, 80, 100, 120, 140, 150㎍/㎖)로 76.97±1.56, 68.15±1.38, 51.85±1.38, 51.85±5.88, 41.74±1.34, 28.18±0.46, 21.31±0.71, 19.87±2.42, 18.38±1.15이었고, 20㎍/㎖에서부터 유의적으로 감소하였다(Fig. 2C).
OJEF의 A375 세포에 대한 cell cycle arrest 유도능을 확인하기 위하여 MUSE™ analyzer를 이용하여 G0/G1, S, G2/M phase의 %를 각각 측정하였다. 농도별(0, 0.1% DMSO, 20, 40, 60, 80, 100㎍/㎖)로 OJEF를 처리하고 6h 배양 후 세포주기를 확인한 histogram 결과, 농도의존적으로 G0/G1 phase peak가 높아지는 것이 관찰되었으며, 반대로 G2/M phase peak는 낮아지는 현상이 관찰되었다(Fig. 3). Fig. 3의 histogram을 %으로 변환한 결과, G0/G1 phase의 경우 44.17±6.71인 control과 비교하여 농도별(20, 40, 60, 80, 100㎍/㎖)로 48.30±4.82, 50.10±7.00, 57.47±3.50, 59.47±4.48, 61.80±6.26이었고, 60㎍/㎖에서부터 유의하게 증가하였다. S기는 전체 농도에서 큰 차이를 보이지 않았으며, G2/M phase인 경우 27.90±5.21인 control과 비교하여 농도별(20, 40, 60, 80, 100㎍/㎖)로 23.63±6.29, 21.13±7.60, 19.10±2.65, 15.83±3.58, 11.43±2.17이었고, 60㎍/㎖에서부터 유의적으로 감소하였다(Table 1).
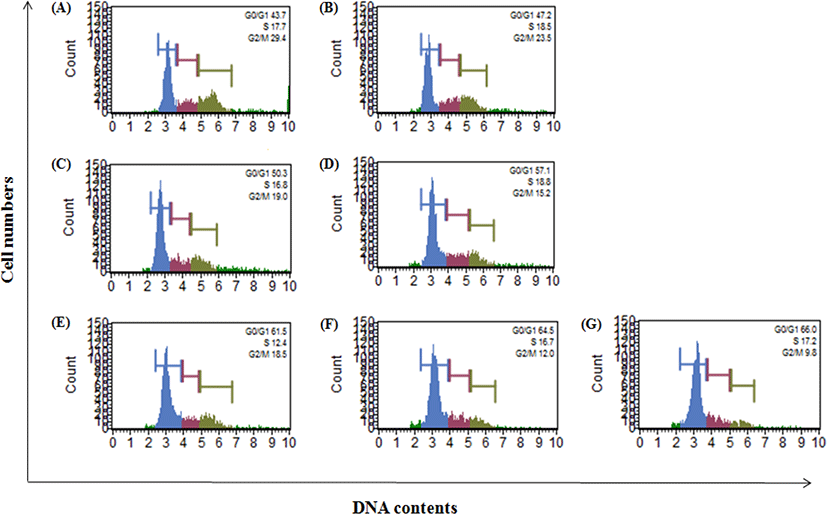
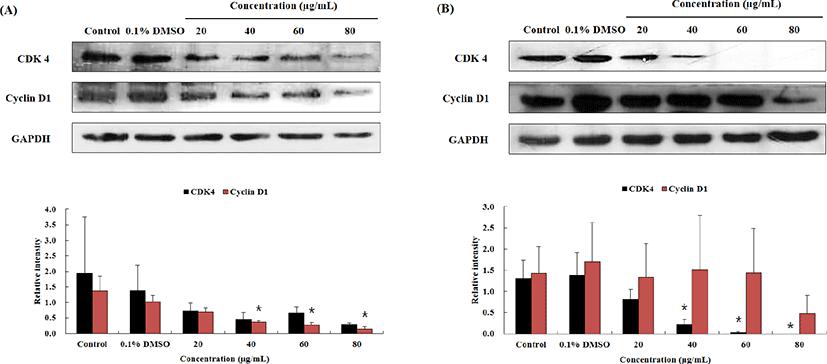
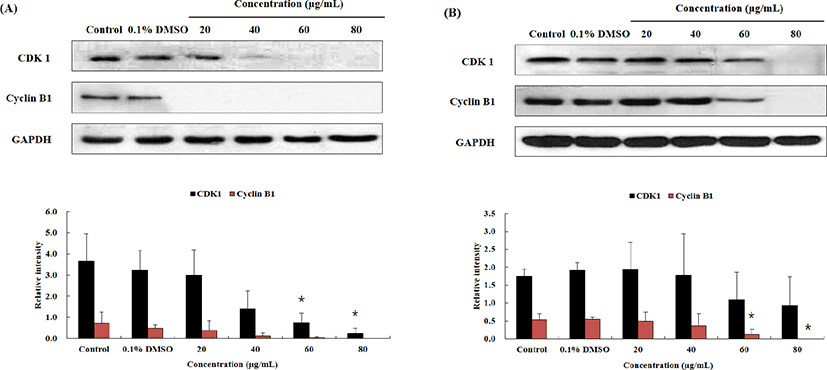
Fig. 3와 Table 1에서 보이듯, OJEF 처리 후, A375 human melanoma cells의 세포증식이 G1 phase arrest에 의해서 세포 증식이 억제되는 것을 확인하였다. 이와 관련하여 각 세포주기를 조절하는 인자에 대하여 단백질 수준에서 western blotting을 통하여 확인하였다. MUSE™ analyzer를 이용하여 실험한 cell cycle 결과에서 G1 phase는 유의적으로 arrest가 증가하였고, G2/M phase으로의 진행은 유의적으로 감소하였다. G1 phase에서는 세포의 크기가 DNA 복제에 적당한지 확인하는 시기인데, 이때 관여하는 단백질은 CDK 4와 cyclin D1 complex 이다. 또한 분열기로 진행하기 전에 DNA 복제가 종결되었는지 확인하는 시기는 G2/M phase로서 관여하는 단백질은 CDK1과 cyclin B1 complex 이다. Fig. 4는 OJEF 처리 후, 농도(0, 0.1% DMSO, 20, 40, 60, 80㎍/㎖) 및 시간(12 및 24h)에 따른 CDK4와 cyclin D1의 발현정도를 확인한 결과로서, 농도 및 시간 의존적으로 단백질 발현량이 감소하는 것을 확인할 수 있었다. 각 농도별 band 강도를 house keeping gene인 GAPDH와 비교하여 수치로 변환한 것을 그래프로 나타낸 결과를 보면, 12h의 CDK 4 경우, 1.93±1.82 인 control과 비교하여 농도별(20, 40, 60, 80㎍/㎖)로 0.73±0.25, 0.46±0.22, 0.66±0.20, 0.29±0.05으로 감소하였고, cyclin D1의 경우, 1.37±0.47인 control과 비교하여 농도별(20, 40, 60, 80㎍/㎖)로 0.70±0.13, 0.37±0.04, 0.29±0.06, 0.14±0.07으로 감소하는 경향을 보였으며, 40㎍/㎖에서부터 유의적으로 감소하였다(Fig. 4A). 그리고 24h의 CDK 4 경우, 1.31±0.43인 control과 비교하여 농도별(20, 40, 60, 80㎍/㎖)로 0.82±0.23, 0.22±0.11, 0.03±0.03, 0으로 감소하는 경향을 보였으며, 40㎍/㎖에서부터 유의적으로 감소하였다. cyclin D1의 경우, 1.43±0.63인 control과 비교하여 농도별(20, 40, 60, 80㎍/㎖)로 1.34±0.79, 1.52±1.27, 1.44±1.05, 0.48±0.42으로 감소하였다(Fig. 4B). CDK 1과 cyclin B1 complex의 경우에서도 농도 및 시간 의존적으로 단백질 발현이 감소하는 것을 확인하였다(Fig. 5). 12h의 CDK 1 경우, 3.66±1.31 인 control과 비교하여 농도별(20, 40, 60, 80㎍/㎖)로 2.99±1.22, 1.41±0.84, 0.76±0.45, 0.24±0.26으로 감소하는 경향을 보였으며, 60㎍/㎖에서부터 유의적으로 감소하였다. cyclin B1의 경우, 0.71±0.53인 control과 비교하여 농도별(20, 40, 60, 80㎍/㎖)로 0.37±0.48, 0.13±0.13, 0.03±0.06, 0으로 감소하였다(Fig. 5A). 그리고 24h의 CDK 1 경우, 1.74±0.22 인 control과 비교하여 농도별(20, 40, 60, 80㎍/㎖)로 1.94±0.76, 1.79±1.14, 1.09±0.77, 0.94±0.80으로 감소하였고, cyclin B1의 경우, 0.54±0.16인 control과 비교하여 농도별(20, 40, 60, 80㎍/㎖)로 0.49±0.26, 0.36±0.34, 0.12±0.15, 0으로 감소하는 경향을 보였 으며, 60㎍/㎖에서부터 유의적으로 감소하였다(Fig. 5B).
Ⅳ. 고 찰
약리성분과 관련하여 와송은 대량의 초산을 함유하고 있다고 하였고20), 와송의 성분에 대한 연구에서 Friedelin, epi-friedlanol, grutinone, glutinol, triterpenid, kaempferol, quercetin, flavonoidβ-sitosterol, cam-pesterol, fatty acid ester, quercetin, flavonoid 등 다양한 약리적 성분이 존재하는 것으로 밝혀졌으며13,21), 박 등22)은 와송의 Triterpene 및 Steroid 성분으로 taraxerone, stigmast-4-3-one and ergost-4-ene-3-one을 분리하였다. 와송의 항암 효능에 대한 관심은 비교적 오래되어 한의학계에서는 1966년에 초보적인 실험이나마 항암작용과 독성에 대한 고찰이 이루어졌으며23), 최근에 와서는 위암11), 대장암19,24), 간암18), 전립선암25), 난소암과 자궁경부암26) 및 백혈병27) 등에서 항암효능이 밝혀져 있다. 그러나 분자생물학적 수준에서의 항암 기작 연구18,19,28)는 부족한 실정이다.
악성 흑색종은 림프절을 통해 전이되고 전체 피부종양에 의한 사망자의 약 80%를 차지하는 악성 피부종양이다29). 멜라닌 세포가 존재하는 어느 부위에나 발생할 수 있으며, 다른 피부암과 달리 성장 속도가 매우 빨라 발생 초기에 다른 장기로 전이율이 높아 악성도가 높은 종양으로 알려져 있다6). 백인들의 경우 미국, 호주, 유럽 등지에서 발생 빈도가 점차 증가하는 추세이며, 과도한 자외선 노출과 유전적인 요인 등이 악성 흑색종의 발병에 중요한 원인이다30). 흑색종이 다른 피부암보다 더 치명적인 이유는 항암화학요법에 대한 저항성을 가지고 있기 때문인데 그 기전은 아직 밝혀지지 않았지만 세포 사멸 기전과 연관되어 있다고 생각되고 있으며, 또한 이것이 질병의 진행과 전이에 연관되어 있다고 알려져 있어 세포 사멸에 관한 기전연구가 절실히 요구된다.
세포주기 조절의 관점에서 암세포는 세포주기의 비정상화에 기인한 질병으로서 특정 시기의 세포주기 억제는 세포주기 checkpoint 각각의 시기에 요구되는 세포주기 조절인자의 발현 및 활성 여부에 따라 조절된다31,32). 첫 번째는 restriction point로써 G1기에 세포크기가 정상인지 DNA 복제에 필요한 물질들을 제대로 합성했는지 검사하는 시기이며, 이 지점에 관여하는 세포주기 유전자는 CDK2/4/6와 cyclin D/E이다33). 두 번째는 G2/M transition으로써 세포분열을 진행하기 전에 DNA 복제가 종결되었는지를 확인하는 지점이며, 이 지점에 관여하는 세포주기 유전자는 CDK1과 cyclin B이다34). 세 번째는 M기 내의 metaphase-anaphase transition으로써 방추사가 제대로 부착했는지 확인하는 지점이다. 세포분열이 일어나기 위해서 반드시 필요한 인자를 mitosis promoting factor (MPF)라고 하며, CDK1과 cyclin B 결합형 효소를 말한다35). 정상 세포에서는 DNA 손상 시 세포 주기가 G1/S기 또는 G2/M기에서 정체되어 손상된 DNA를 수복할 수 있는 DNA 수복 기전이 존재한다. 그러나 과다한 DNA 손상이 야기되어 DNA 수복이 어려워지게 되면 G1/S 또는 G2/M check point에서 이를 감지하여 apoptosis가 유도되어 게놈상의 안전성을 유지하고 있다36). 본 연구에서는 OJEF 처리 시 농도 의존적으로 G1 phase는 유의적으로 arrest가 증가하였고, G2/M phase으로의 진행은 유의적으로 감소하였다. OJEF를 농도별로 처리하고 6h 배양했을 때 100㎍/㎖ 처리군에서 control에 비해 G0/G1 phase peak의 %는 1.4배 증가 하였고, 같은 농도에서 G2/M phase peak의 %는 control에 비해 2.44배 감소하였다. 이와 같은 결과를 통하여 OJEF가 A375 human melanoma cells의 G1 phase를 arrest 시킴으로서 암세포 증식을 억제함을 확인하였다. 이는 G1기 이행단계를 조절하는 CDK4와 cyclin D1의 단백질 발현량이 농도 및 시간 의존적으로 감소한 것과 깊은 관련이 있을 것으로 판단된다. 또한 CDK 1과 cyclin B1가 농도 및 시간 의존적으로 감소한 것은 G2/M phase 로의 진행을 늦추는 것과 연관된 것으로 보인다. 이를 통하여 OJEF가 A375 흑색종 세포의 세포주기 정지를 일으켜 세포분열이 제어됨으로써 증식이 억제될 것으로 사료된다.
이상의 결과를 종합하면 OJEF는 흑색종 세포주인 A375 cells에 선택적으로 시간 및 농도 의존적인 세포독성을 보였으며 세포주기 조절과 관련된 CDK1, cyclin B1, CDK4, cyclin D 억제로 G1 phase와 G2/M phase에서 세포주기 정지가 일어나 세포분열이 제어되었음을 확인하였다. 따라서 와송 유래 생리 활성 물질의 흑색종에 대한 항암 효과가 확인되었으므로 앞으로 와송 추출물의 항암제로의 개발을 위해서는 세포자멸사에 대하여 보다 정확한 신호전달 기전 연구와 in vivo 연구가 필요하리라 사료되며 이를 바탕으로 다양한 질병으로 고통 받는 환자들을 치료하는데 도움이 될 치료제로서 개발 가능성이 매우 높다고 사료된다.
Ⅴ. 결 론
A375 인체 흑색종 세포에 대한 와송의 항암효과를 실험적으로 규명하기 위하여 OJEF를 투여한 후 암세포 증식 억제효과와 cell cycle arrest 유도능을 측정하고 western blotting을 통하여 apoptosis 관련 유전자 및 단백질의 발현을 측정하여 다음과 같은 결과를 얻었다. A375 인체 흑색종 세포의 증식이 OJEF의 처리농도와 배양시간에 의존적으로 감소하는 것을 알 수 있었고 세포주기 조절을 보면 농도 의존적으로 G1 phase는 유의적으로 arrest가 증가하였고, G2/M phase으로의 진행은 유의적으로 감소하였다. CDK4와 cyclin D1, CDK 1과 cyclin B1의 단백질 발현량이 농도 및 시간 의존적으로 감소하였다.