



Ⅰ. 서 론
자궁암은 암으로 인한 사망 중 가장 중요한 요인 중 하나이며1)인체 유두종 바이러스(HPV)가 자궁암의 중요원인을 제공하고 있다2). 다른 위험한 요소들은 다수의 성교 대상자, 어린 나이의 성교, 흡연 등을 포함하고 있으며 이는 성교를 통해 옮겨진다는 것이 확실하다3).
Amygdalin은 많은 nitrilosides 계통 물질의 하나이며 이 물질은 아몬드, 살구 및 다른 장미과 식물들에 많이 들어있다. Amygdalin은 비타민 B17로도 알려져 있으며, 이전의 연구들은 D-Amygdalin이 종양치료에 효과가 있다는 것을 보여주고 있다4).
세포사멸은 세포 예정사라는 생리학적 또는 항상성 역학 구조 역할을 수행하며5) 암, 자기면역의 기능장애, 후천성 면역결핍증(AIDS), 신경퇴행성 질병 등의 발병 및 병리생리학에 밀접하게 연루되어 있다6). 세포사멸은 여러 종양 진행 단계들에 상당한 영향을 끼치는 것으로 알려졌으며7), 종양 발전을 차단하는 핵심적 역할을 수행하는 것으로도 알려져 있다8).
이 세포사멸의 경로에 비정상적인 활성 혹은 비활성화는 결국 발암현상으로 이어지는 통제 불능의 세포 분열 증식을 유도 할 수도 있다9). 종양세포 내부의 어떤 균에 의한 세포사멸의 유도는 유용한 암 치료 방법으로 인식 되고 있다10).
세포사멸의 영향을 받고 있는 세포들은 염색체 응축, 핵 무사분열, 세포사멸체 형성 같은 여러 형태학적 특징을 나타낸다11). 세포사멸의 진행이 몇몇 단백질의 유전자 합성에 의해 규제 된다는 것은 기록으로 잘 나타나 있으며 Bcl-2군은 각기 수행하는 역할에 의해 anti-apoptotic 단백질, pro-apoptotic 단백질 2가지로 구분된다.
Bcl-2 중 anti-apoptotic 단백질은 apoptotic 경로를 규제하고 또한 세포사멸에 대항해 보호하기도 한다는 것으로 알려져 있는 반면 pro-apoptotic 단백질 중 Bax는 세포사멸 과정에서 선별적이면서도 매우 많은 양의 유전자가 합성되어지고 세포사멸을 촉진시키는 것으로 알려져 있다12). Bcl-2 단백질에 대한 Bax 단백질의 비정상적인 비율은 백혈병, 유방암, 폐암 등의 다양한 암에서 관찰되었다13-15).
세포사멸을 유도하는 항암 약품들의 분자적 구조는 세포사멸의 진행에서 Caspases의 활성화에서 나타나는 일련의 반응들 중에서 확인해야할 중요한 항목인 cytochrome c 방출과 관계가 있으며16-18) Cysteine 단백분해효소 부류인 Caspases는 apoptotic 진행에 핵심적인 역할을 하는 것과 함께 포유류 세포의 단백질 가수분해 난할에서 나타나는 일련의 반응들을 유발하는 것으로 알려져 있다. 세포사멸의 중요한 역할자중 하나이며 Caspases군 중 가장 광범위하게 연구된 Caspase-3의 특징은 다양한 단백질 가수분해 난할 과정의 전체, 혹은 일부를 책임지는 것이다19).
이에 저자는 杏仁으로부터 추출한 Amygdalin의 자궁경부암세포 ME-180에 대한 항암 효과를 실험하기 위해 세포 생존율 측정, TUNEL 측정, DAPI 염색, flow cytometric analysis, DNA fragmentation 측정, western blot 그리고 Caspase-3 효소 활성 측정을 통하여 유의한 결과를 얻었기에 보고하는 바이다.
Ⅱ. 실 험
껍질을 벗겨낸 杏仁 500g과 4%의 구연산이 담긴 10ℓ 용액을 2시간동안 환류 시킨다. 여과한 후 열이 식지 않았을 때 HP-20으로 포장된 column을 통과시켰다. column 내부에서 흡수된 물질을 에탄올로 희석시킨 후에 농축시켰다. 4.2g의 Amygdalin(0.84%의 농도)은 에탄올로 재결정체화 시킴으로서 차단되고 Amygdalin은 high-pressure liquid chromatography (HPLC;shiseido, Tokyo, Japan)로 순도 99.0%인 것을 사용하였다.
3-(4,5-dimenthylthiazol-2-yl)-2, 5-diphenylterazolium bromide(MTT)측정 kit는 Boehringer Mannheim (Mannheim, Germany)사, DNA fragmentation 측정 kit는 TaKaRa (Shiga, Tokyo, Japan)사, Caspase-3 측정 kit는 CLNTECH (Palo Alto CA, USA)사에서 구입하였다. 3,3-diaminobenzidine(DAB), 4,6-diamidino-2-phenylindole (DAPI), Propidium iodide(PI)와 paraformaldehyde(PFA) 그리고 다른 약물들은 Sigma Chemical (St.Luis, Mo, USA)사로부터 구매하였다.
ME-180 자궁경부암세포는 한국세포은행(KCLB, Seoul, Korea)으로부터 구입하였다. 10%의 heat inactivated fetal bovine serum을 포함한 RPMI 1640 media (Gibco BRL, Grand Island, NY, USA)로 배양기(37°C, 5% CO2, 95% O2)에서 배양하였으며 media들은 2일에 한번씩 교환하였다.
인체의 ME-180 자궁암세포들은 96-well plate에서 well 당 100㎕에서 배양하였고 Amygdalin의 세포 특성을 알아보기 위해 세포들을 1㎍/㎖, 10㎍/㎖, 100㎍/㎖, 1㎎/㎖ 및 10㎎/㎖ 농도에서 Amygdalin을 24시간 동안 처리하였으며 대조군의 세포들은 처리하지 않은 채 놔두었다. Phosphate -buffered saline(PBS) 내 5㎎/㎖의 MTT를 함유하고 있는 10㎖의 MTT label 시약을 각각의 well에 첨가 하여 4시간 동안 배양하였다. 0.01M hydrochloric acid(HCI)에서 10%의 sodium dodecyl sulfate를 함유하고 있는 100㎕의 가용성 용액을 각각의 well에 첨가한 후 세포들을 다시 12시간 동안 배양하였다. 이 후 microtiter plate reader (Bio-Tek, Winooski, VT, USA)로 690nm 기준파장에서 595㎚ 실험파장으로 흡광도를 측정하였다. 광학밀도(O.D)는 기준파장과 실험파장에서 관측한 파장의 차이로 계산하였으며 생존 능력의 백분율은 (약물 처리 견본O.D / 대조군O.D)×100으로 계산하였다.
apoptotic 세포의 in situ 검출을 위해 peroxidase in situ 검출 kit 내에 있는 ApoTag를 사용하여 측정하였다. 챔버당 2×104 세포 밀도로 4개의 챔버 슬라이드에서 ME-180 세포를 배양하였다. 1㎎/㎖과 10㎎/㎖ 농도의 Amygdalin을 ME-180 인체 자궁암세포에 24시간 노출시킨 후에 세포들을 PBS로 세척하고 4℃에서 10분간 4%의 PFA에 고정시켰다. 고정된 세포들은 가습 환경 37℃에서 60분간 terminal deoxynucleotidyl transferase(TDT) 내에서 digoxigenin -conjugated dUTP로 배양하였다. 실온에서 약 10분간 stop/wash 완충액에 넣었다. 세포들은 peroxidase와 복합된 anti-dioxygenin antibody에서 30분간 배양하였으며 DNA단편들은 DAB를 peroxidase에 사용하여 억제시켰다.
인체의 자궁암세포 ME-180에서 세포사멸이 Amygdalin에 의해 유도되는가를 결정하기 위해 DAPI 염색을 실시하였다. 4개의 챔버 슬라이드에서 세포를 배양하여 1㎎/㎖와 10㎎/㎖ Amygdalin 용액에서 ME-180 세포에 24시간 동안 처리한 다음 PBS로 두 번 세척하고, 4% PFA에서 30분간 배양하여 고정한 다음 PBS로 두 번 세척하였다. 어둠 속에서 30분간 1㎍/㎖의 용액에서 배양 후 세포들을 형광 현미경(Zeiss, Oberkochen, Germany)으로 관찰 하였다.
1㎎/㎖와 10㎎/㎖ Amygdalin을 인체 자궁암세포 ME-180에 24시간 처리한 후에 세포들을 수집하여 1시간동안 -20℃에서 75%의 에탄올이 있는 PBS에 고정시키고 100㎍/㎖의 RNase로 배양하여 PBS내에 20㎍/㎖ propidium 요오드 상태에서 염색하였다. 염색된 세포들은 37℃에서 30분간 배양하고 FACScan(Becton Dickinson, San Jose, CA, USA)으로 분석 하였다.
Apoptotic DNA의 분할 검출을 위해 ApopLadder EXTM DNA 무사분열 측정kit를 이용하여 DNA fragmentation 측정을 실시하였다. 인체 자궁암세포 ME-180을 수집하여 1㎎/㎖와 10㎎/㎖ Amygdalin을 24시간 처리 하고 PBS로 세척하였다. 세포들은 100㎕의 용해액으로 용해하였고 용해 분리물은 37℃에서 1시간 동안 10㎕ 효소 B에서 처리한 후 56℃에서 10㎕의 효소 A를 함유하고 있는 10㎕의 10% SDS 에서 배양 하였다. 이 혼합물에 500㎕ 에탄올과 70㎕ 침전제를 첨가한 후 약 15분 동안 원심분리 처리하였다. 이후 에탄올의 합성 pellet 세척으로 DNA를 추출하고 이들을 Tris-EDTA (TE)용해액에 부유시켰다. DNA Fragmentation은 ethidium bromide를 함유한 2% agarose gel 전기 이동에 의해 시각화하였다.
인체 자궁경부암세포 ME-180에 1㎎/㎖와 10㎎/㎖ Amygdalin을 24시간 처리한 후에 세포들을 50mM HEPES(pH 7.5) 150mM Nacl, 10% glycerol, 1% Triton X-100, 1.5mM magnesium chloride hexahydrate, 1mM ethyleneglycol-bis-(β-aminoethyl ether)-N, N'-tetraacetic acid(EGTA), 1mM phenylmethylsulfonyl fluoride(PMSF), 2㎎/㎖ leupeptin, 1㎎/㎖ pepstatin, 1mM sodium ortho vanadate 그리고 100mM sodium floride를 함유하고 있는 얼음 냉각 세포 용해액에 용해시켰다. 이후 혼합물질은 30분간 4℃에서 배양하였다. 상층액의 급속냉동에 따른 마이크로 원심분리에 의해 세포 잔해를 제거하였다. 단백질 응축액들을 Bio-Red colorimetric protein 측정kit로 측정하였다. 단백질 30㎎을 SDS-polyacrylamide gel에 분리시켰고 nitrocellulose membrane에 이전시켰다 (Schleicher & Schull GmbH, Dassel, Gremany).
Mouse Bax 항체(Santa Cruz Biotech, Santa Cruz, CA, USA)와 mouse Bcl-2 항체(Santa Cruz Biotech)를 초기 항체로 사용 하였고 Horseradish peroxidase와 Bax, Bcl-2의 anti-mouse 항체(Amersham Pharmacia Biotech, Freiburg, Germany)를 복합해 2차 항체로 사용하였다. Band 검출은 enhanced chemiluminescene(ECL)검출 시스템(Santa Cruz Biotech)을 사용하였다.
Caspase 활성은 Caspase-3 측정kit를 제조회사의 첨부서에 따라 ApoAlert를 사용하여 측정하였다. Amygdalin을 인체 자궁경부암세포 ME-180에 1㎎/㎖ 와 10㎎/㎖의 농도로 24시간 처리한 후 50㎕의 냉장된 세포 용해액에서 용해 시켰다. 각각의 용해물질을 50㎕의 1/2 반응액(dithioreitol 함유)에 첨가하고 알맞게 복합된 5㎕의 기질을 1mM의 용액으로 첨가하였다. 그리고 혼합물을 37℃에서 1시간 동안 중탕하면서 배양하였다. 흡광도는 microtiter를 405㎚의 실험 파장으로 하여 측정하였다.
인체 자궁경부암 세포 ME-180에 1㎎/㎖ 와 10㎎/㎖ Amygdalin을 24시간 동안 처리한 후 세포들을 500㎕ cytofix/cytoperm (PharMingen, San Diegio, CA, USA)에 재부유시키고 얼음 위에서 20분간 배양하였다. 이후 500㎕ perm/wash buffer(PharMingen)으로 세척하고 세포들을 100㎕ perm/wash buffer에 염색하기위해 재부유시켰다. 정상 rabbit IgG(Santa Cruz Biotechnology, Santa Cruz, CA, USA)를 얼음 위에서 20분간 첨가시켰다. Phycoerythrin(PE) -복합 polyclonal rabbitti anti-active Caspase-3 항체(PharMingen) 20㎕을 첨가시켰다. 이후 세포들을 어둠속에서 30분간 얼음 위에서 배양하고 500㎕ perm/wash buffer로 두 번 세척 한 후 500㎕ 염색용해액에 재부유시키고, FACScan(Becton Dickinson)을 이용하여 분석 하였다.
Ⅲ. 결 과
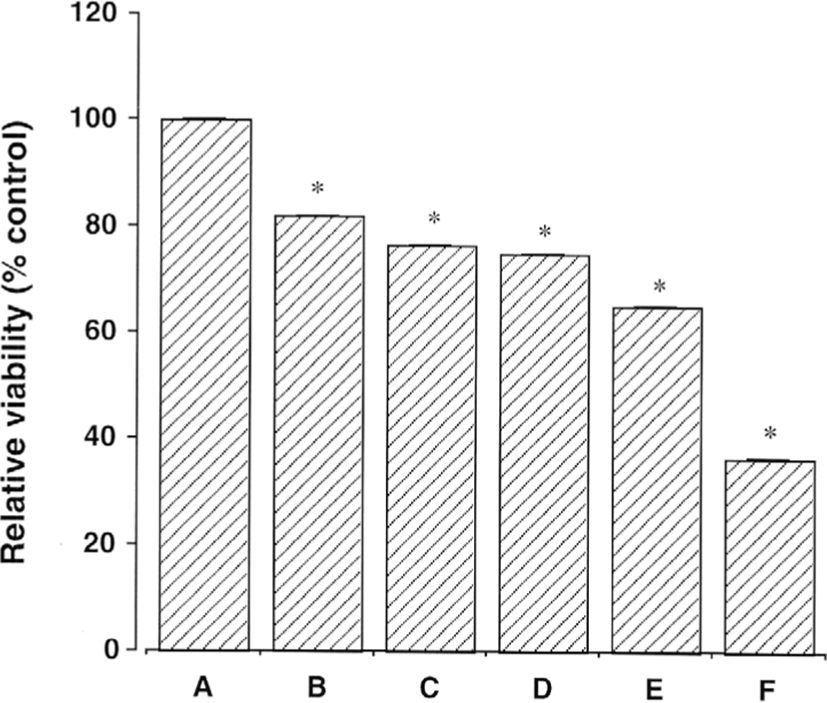
Fig. 1에서 보이는 것과 같이 1㎍/㎖, 10㎍/㎖, 100㎍/㎖, 1㎎/㎖ 그리고 10㎎/㎖ Amygdalin을 24시간 배양 했을 때 세포 생존율은 대조군 100.00 ± 1.19%에 비해 비교한 값은 각각 81.33 ± 0.70%, 76.61 ± 1.78%, 74.79 ± 1.62%, 65.22 ± 0.71%, 그리고 36.37 ± 0.49% 대조 표준값이었다. Amygdalin 농도의 증가에 따른 세포 독성이 증가하는 경향을 나타내었다. 이런 결과에 기초하여 진행된 실험에서 세포독성을 유도하는 Amygdalin의 특성을 나타내는 Amygdalin의 농도로 1㎎/㎖와 10㎎/㎖을 선택하였다.
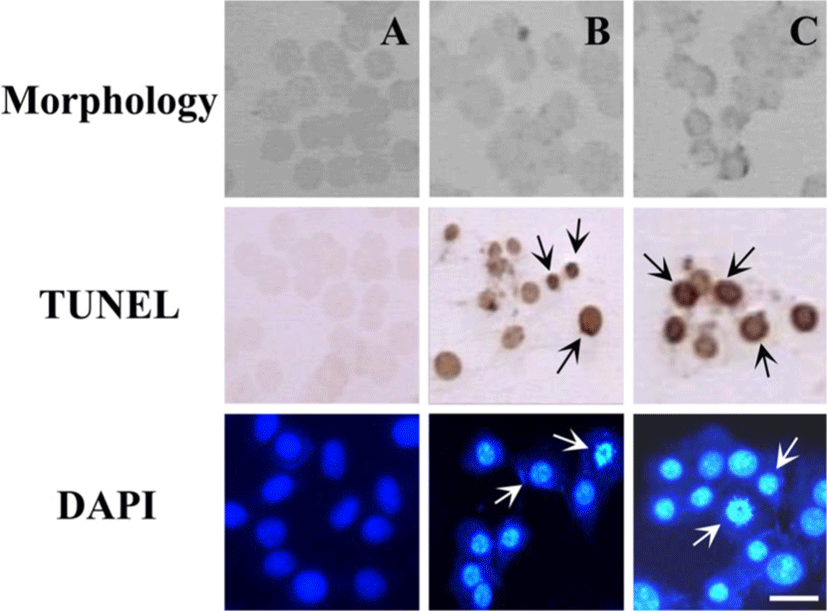
세포 형태에 미치는 Amygdalin의 효과를 위상차 현미경을 사용하여 관찰 하였다. Fig. 2에서 보이는 것과 같이 1㎎/㎖와 10㎎/㎖ 농도의 Amygdalin으로 24시간 동안 배양한 세포들은 세포 수축, 세포질 응결, 그리고 불규칙함을 보였다. 이러한 형태학적 특징들은 Amygdalin이 ME-180 세포에서 세포사멸을 유도한다는 것을 보여준다. Amygdalin에 의해 ME-180 세포에서 세포사멸이 유도된다는 것을 보다 더 확인하기 위해 Amygdalin 처리된 세포들을 DAPI 염색과 TUNEL 측정을 하여 생화학적으로 분석하였다. Fig. 2에서 보이는 것과 같이 TUNEL 양성 세포들은 밝은 현미경 아래에서 dark brown의 얼룩이 졌으며 핵응결을 보인 TUNEL 양성 세포들은 1㎎/㎖과 10㎎/㎖ 농도로 24시간 Amygdalin 처리된 세포에서 관찰되었다. DAPI 염색에서, 특별히 핵을 염색하는 DAPI 처리를 한 후 형광 현미경을 통해 세포들을 관찰하였고, 핵응결 존재, DNA framentation 그리고 Amygdalin 처리 세포에서 핵주위의 세포사멸을 밝혀내는 측정을 했다. 이러한 결과를 통해 Amygdalin이 인체 자궁암세포 ME-180에서 세포사멸 형태적 변화를 유도한다는 것을 보여주고 있다.
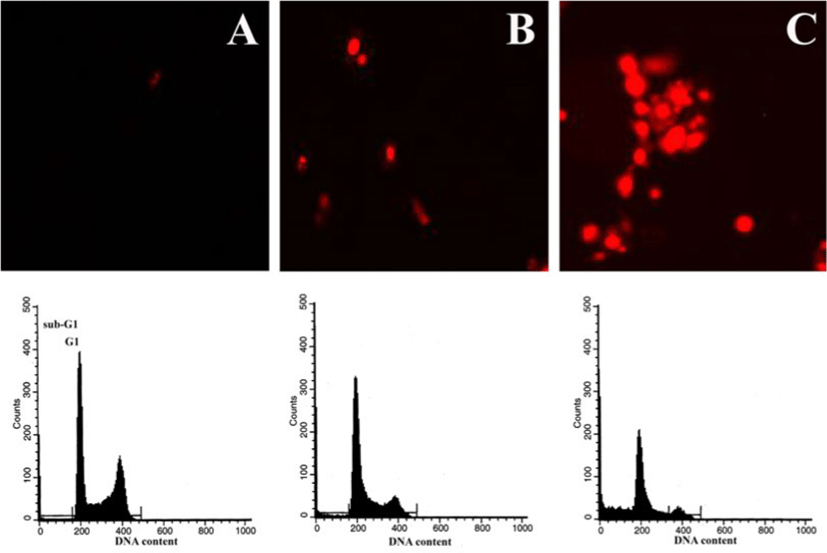
Flow cytometric analysis는 세포들을 1㎎/㎖와 10㎎/㎖ Amygdalin으로 24시간 동안 처리한 후, sub-G1 반응기에서 세포 파편들이 대조군 2.14% 보다 18.69% 그리고 49.88%로 각각 증가되는 것을 보여준다(Fig. 3). 이러한 결과들은 인체 자궁암세포 ME-180에서 Amygdalin이 세포사멸을 유도한다는 것을 보여준다.
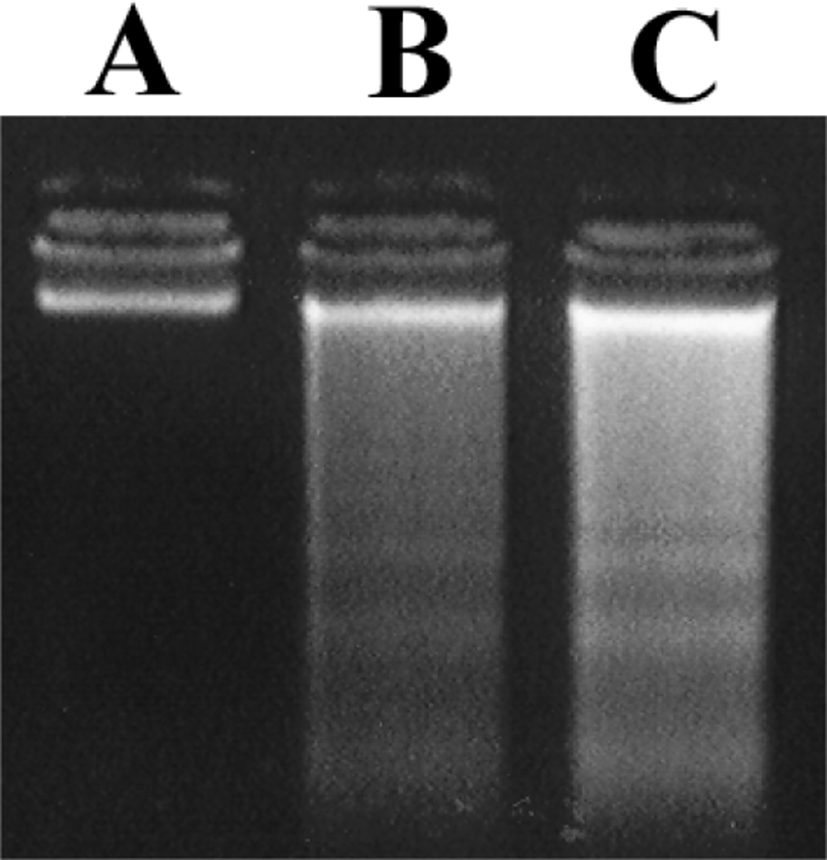
Amygdalin에 의한 세포사멸 유도를 확인하기 위하여 endonuclease 활성을 반영하는 DNA fragmentation을 측정하였다. ME-180 암세포를 24시간 동안 1㎎/㎖와 10㎎/㎖ Amygdalin 처리하면 특징적 사다리 무늬의 명확한 fragmentation이 나타남을 electrophoresis를 통해 알 수 있다(Fig. 4).
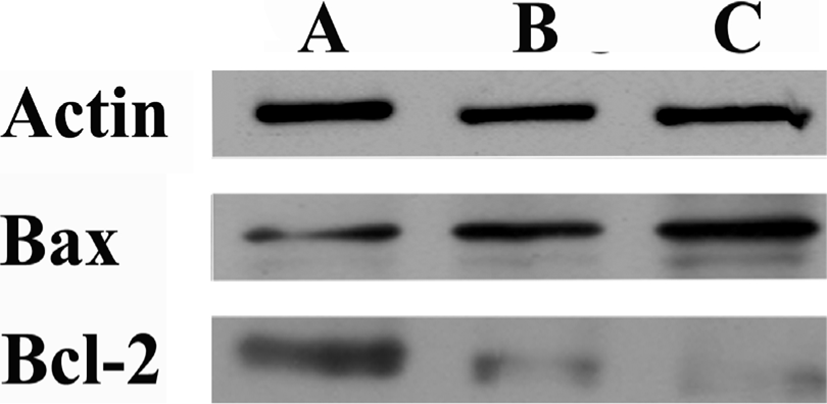
Amygdalin이 Bax 단백질과 Bcl-2 단백질의 합성에 끼치는 영향을 조사하였다. ME-180 암세포에 1㎎/㎖와 10㎎/㎖ 24시간 Amygdalin 처리한 후, Bax 단백질(24kDa)의 합성은 증가 되었고 이와는 반대로 Bcl-2단백질(25kDa)의 합성은 감소되었다(Fig. 5). 이러한 결과는 Amygdalin이 Bax단백질의 합성은 높이고 Bcl-2 단백질의 합성은 억제함을 보여준다.
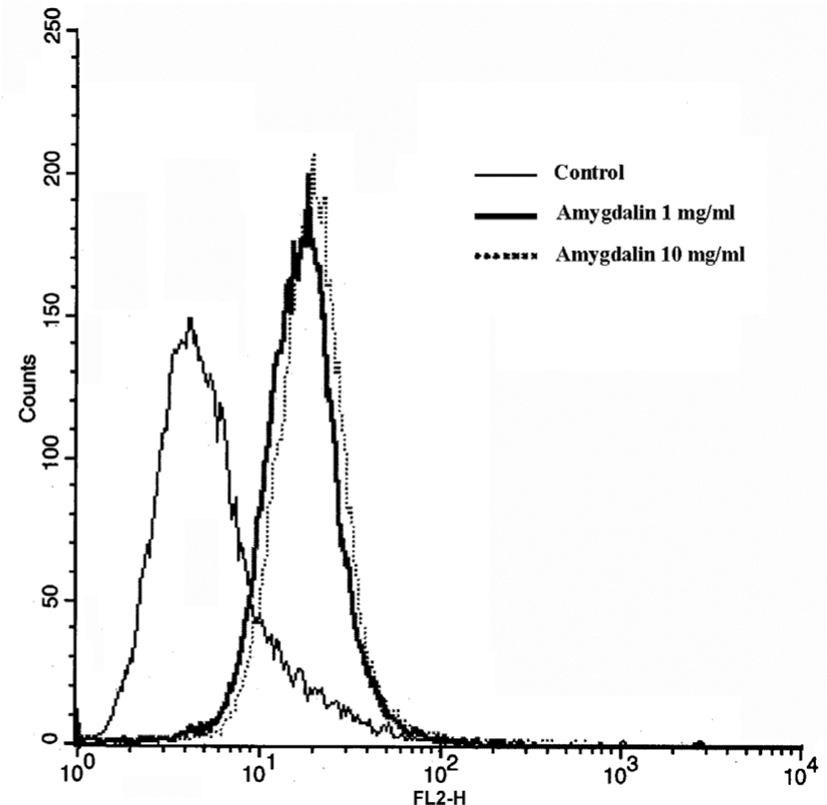
1㎎/㎖와 10㎎/㎖ Amygdalin 처리를 24시간 한 후 Active Caspase-3 합성이 대조군 5.27% 보다 34.5% 그리고 48.5%로 각각 증가 하였다(Fig. 6).
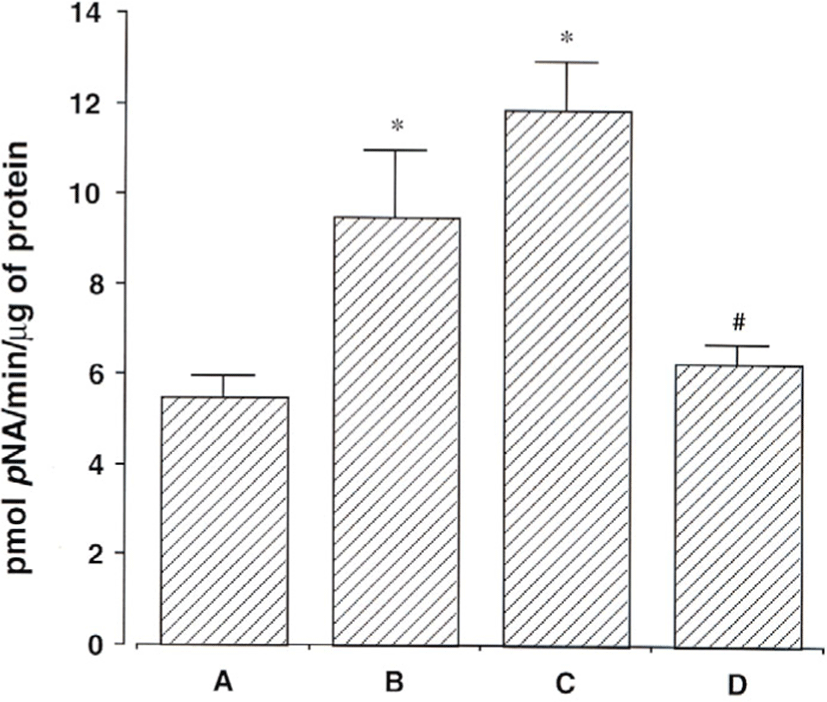
Caspase-3 효소 활성은 DEVD peptide-nitroannilide(pNA)를 사용하여 측정하였다. Amygdalin을 1㎎/㎖와 10㎎/㎖에 24시간 동안 노출 시킨 후, 효소 활성을 해 본 결과 DEVD-pNA 분할비율이 대조군 5.53±0.46pmol)에서 9.50±1.48pmol과 9.50±1.48pmol로 각각 증가함을 알 수 있었다(Fig. 7).
Ⅳ. 고 찰
杏仁의 주요성분인 Amygdalin은 자체의 항암 활성이 수년 동안 연구되어져 왔다20-22). 최근에는 D-Amygdalin이 항암 시약으로서의 가능성으로 인해 주목 받고 있다23,24). D-Amygdalin의 부분입체 이성질체인 neoamygdalin (L-mandelonitrile-b-D-genetiobioside)은 암 치료에 대하여 비효과적이어서 암 치료제로서 Amygdalin을 사용할 때 분리해내야 한다25). Syrigos 등22)은 Amygdalin이 항체유도 효소 nitrile 요법을 통해 종양 치료 요법에 사용될 수 있다고 보고 하였다. Amygdalin은 암 치료요법에서 상당한 논쟁의 주제로서 다루어져 왔다20-26).
세포사멸은 암세포에 대항하는 보호기능을 제공한다27). 세포사멸의 유도는 암조절을 위한 치료 전략에 이상적인 방법이다28,29). 본 연구의 목적은 Amygdalin이 인체 자궁암세포 ME-180의 사멸을 유도하느냐를 밝혀내는 것이며 동시에 Amygdalin이 자궁암 치료약으로 사용될 수 있는지를 밝혀내는 것이다.
세포 생존율 측정에 의한 세포 독성 분석은 Amygdalin이 자체의 세포 독성 효과에서 농도 의존적이라는 것이 확인 되었고 이러한 세포독성은 1㎎/㎖ 와10㎎/㎖ 농도에서 명확하게 나타났다. PI를 이용한 DNA 용량의 flow cytometric 분석으로부터 초기 sub-G1 단계에서 세포 단편들이 증가한다는 것이 관찰되었다. ME-180세포에 Amygdalin 처리 이후에 세포순환의 다양한 파편들 사이에서 DNA용량의 분배가 세포사멸의 존재를 암시하는 것 같았다.
세포사멸의 존재를 더욱 증명하는 것으로 DNA fragmentation이 agarose gel electrophoresis를 통해 명확하게 검출 된 것을 들 수 있다. 세포사멸은 endonuclease의 활성화와 연계되어 있으며 또한 electrophoresis 실험에서 사다리 형태 특징을 보인 윤곽이 뚜렷한 단편에서 DNA fragmentation을 초래하는 것도 알려져 있다30,31).
DNA strand가 세포사멸과정에서 발생하고 DNA분자의 nick이 TUNEL 측정을 통해 검출 될 수 있다는 것도 알려져 있다32). 상기 언급된 효과에 부가해서 Amygdalin은 세포의 형태학에서 두드러진 변화를 발생시키는 것으로 알려져 있다. 세포사멸의 엄중한 형태학적 기준들인 이러한 변화들은 DAPI 염색에서 확인 되었다.
세포사멸체는 특징적으로 DAPI로 염색된 Amygdalin 처리세포에서 나타났다. 세포사멸을 겪은 세포들은 세포질 수포, 핵수축, 염색질 농축, 형태의 불규칙성, 진행의 수축 등을 나타낸다고 보고되었다.19).
단백질의 Bcl-2계 군들은 Bax와 자체의 homodimers로 heterodimers의 복잡한 결합을 구성할 수 있는 특징이 있다33). Bax가 세포에서 과다하게 합성 되었을 때, 세포 죽음에 대한 반응으로 세포사멸이 가속화된다. Bcl-2가 과다하게 합성되었을 때에는 Bax 와 같이 heterodimerizes되고 세포사멸은 억제된다. 유추해보면 Bcl-2에 대한 Bax의 비율은 세포사멸에 대한 세포의 감수율 결정인자의 역할을 한다34).
Bax의 과다 합성은 전립선암, 결장암, 자궁암 그리고 난소암등 광범위한 분야의 암세포에서 세포사멸을 유도하는 것으로 알려져 있다35-39). 몇몇 연구에서, 결장암, 유방암 그리고 난소암에서는 Bcl-2 합성이 높은 수치를 보여주고 있음을 설명하고 있다40-42). 본 실험 결과에서 Amygdalin 처리가 Bax 합성을 증가시키고 Bcl-2의 합성을 감소를 초래한다는 것을 알 수 있다.
Cysteine protease 계인 Caspases는 apoptotic 경로의 통합분야이다. Caspases는 apoptotic 진행의 효과기와 DNA fragmentation이 핵 형태학적 변화가 Caspase 활성의 근저로 자리매김한 만큼 간주되어 지고 있다43,44).
특히 Caspase-3은 많은 세포질 표적을 가지고 있으며 세포사멸의 특이한 형태적 특징을 생산한다. 본 실험 결과는 Active Caspase-3 합성의 증가와 Amygdalin에 노출된 세포들 내에서의 Caspase-3의 활성 증가도 밝혀냈다.
위의 결과로 보아 Amygdalin이 자궁암세포 ME-180의 세포사멸을 유도한다는 것을 보여주었고, 인체 자궁암에 항암 효과를 발휘할 수도 있다는 가능성을 제시하였다.
Ⅴ. 결 론
Amygdalin의 자궁경부암세포 ME-180에 대한 항암 효과를 실험하기 위해 세포 생존율 측정, TUNEL 측정, DAPI 염색, flow cytometric analysis, DNA fragmentation 측정, western blot 그리고 Caspase-3 효소 활성 측정을 통하여 다음과 같은 결과를 얻었다.
-
세포 생존율 측정에서 Amygdalin 투여군에서는 대조군에 대해 농도 의존적으로 유의한 감소를 나타냈다.
-
TUNEL 측정을 통한 생화학적 분석에서 Amygdalin으로 24시간 동안 배양한 세포들은 세포 수축, 세포질 응결, 그리고 불규칙함을 나타냈다.
-
DAPI 염색을 통한 세포관찰에서 핵응결 존재, DNA framentation 그리고 Amygdalin 처리 세포에서 핵주위의 세포사멸을 나타냈다.
-
Flow cytometric analysis를 통해 sub-G1 반응기에서 Amygdalin 1㎎/㎖와 10㎎/㎖을 투여한 군이 대조군에 비해 세포 파편들이 증가됨을 알 수 있었다.
-
DNA fragmentation을 통해 특징적 사다리 무늬의 명확한 fragmentation이 나타남을 electrophoresis를 통해 알 수 있었다.
-
Western blot에서 Amygdalin이 Bax 단백질의 합성은 높이고 Bcl-2 단백질의 합성은 억제함을 알 수 있었다.
-
Active Caspase-3 합성에 대해 Amygdalin 1㎎/㎖와 10㎎/㎖을 투여한 군이 대조군에 비해 유의한 증가를 나타냈다.
-
Caspase-3 효소 활성 측정한 바 Amygdalin 1㎎/㎖와 10㎎/㎖을 투여한 군이 대조군에 비해 유의한 증가를 나타냈다.
위의 결과로 보아 杏仁에서 추출한 Amygdalin이 자궁암세포 ME-180의 세포사멸을 유도한다는 것을 알 수 있었으며, 인체 자궁경부암에 대한 항암 효과가 있다고 생각된다.